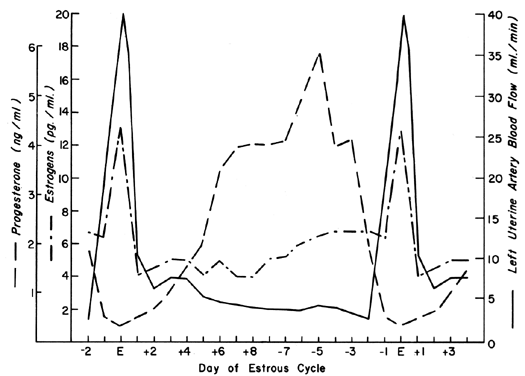
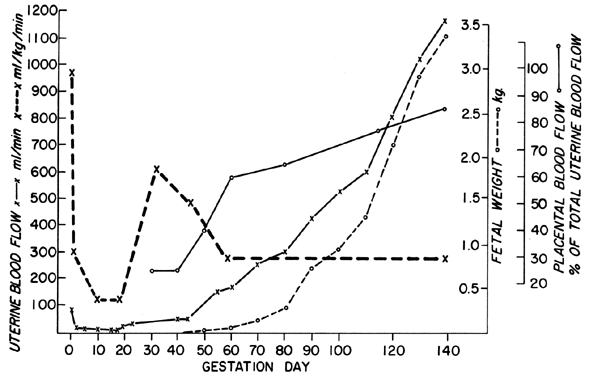
The uterine circulation is exquisitely sensitive to estrogen stimulation, responding with a degree of vasodilatation unparalleled by any other organ of the body to any other stimulus.2 During the ovarian cycle, repetitive patterns of UBF occur reflecting the effect of estrogen secretion and the modulating effects of progesterone. These cyclic patterns are demonstrated most dramatically in animals with short preovulatory cycle phases, such as the cow, sow, and ewe, and are illustrated in Figure 1 with peripheral levels of estrogen and progesterone.3, 4, 5 Following conception in these species, UBF patterns are similar to those in nonpregnant animals until interdigitation of maternal and fetal tissues occurs 17 to 28 days later; then, there is a definitive and progressive increase in UBF that continues until term pregnancy. Since erosion of endometrium by fetal trophoblast occurs much earlier after conception in the primate, it is reasonable to presume that UBF increases earlier in these species, although such responses have never been observed. The overall patterns of total UBF, UBF per unit weight of uterus and its contents, fraction of UBF supplying the placenta, and fetal weight throughout ovine pregnancy are illustrated in Figure 2. The period between 17 and 70 days of gestation is associated with the most rapid changes in UBF and PBF and corresponds with the time of definitive placentation in this species. By mechanisms incompletely understood, blood vessels supplying the placenta progressively dilate during this time, simultaneously causing an absolute increase in total UBF and shunting of this blood from nonplacental to placental tissues.* With subsequent placental maturation, the fraction of UBF supplying the placenta and the absolute amount of PBF increase, but at rates more similar to that of fetal growth; thus, during the last half of pregnancy, UBF per unit weight of uterus and its contents is essentially constant. During this same time, the amount of oxygen extracted from each milliliter of blood is constant and therefore the amount of oxygen delivered per given weight of gravid uterus is also constant.6 Correlative studies in women indicate similar homeostatic patterns from 10 to 40 weeks of gestation and similar oxygen consumption (approximately 10 ml/kg).7 Since women deliver the same amount of oxygen at lower UBF rates than the ewe (~150 versus ~270 ml/kg), it follows that the human placenta is more efficient in this regard. Thus, for fetuses of similar weight, less absolute UBF is required and the pregnancy load on the heart is diminished.
|
|
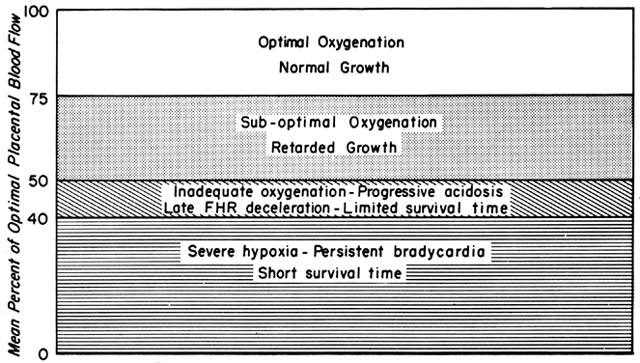
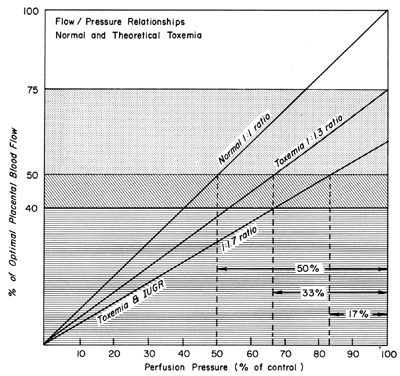
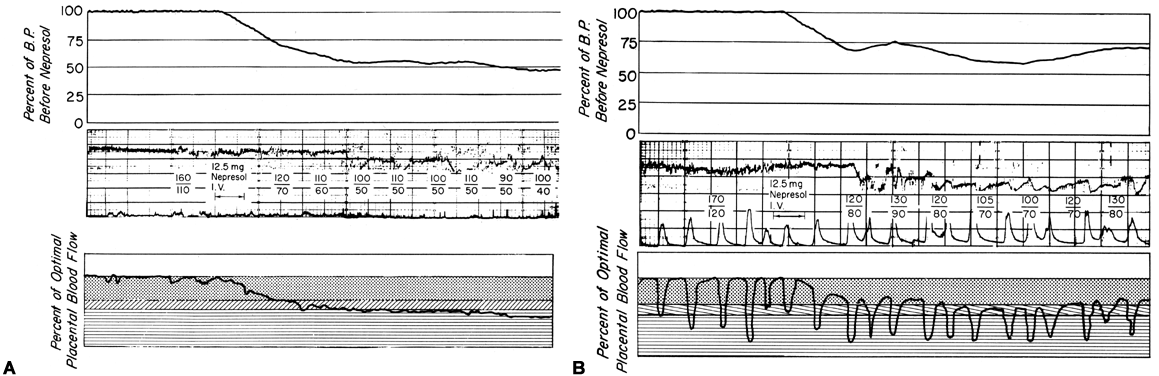
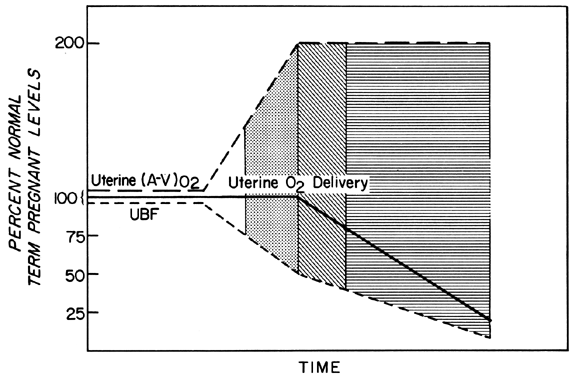
Changes in PBF are accomplished by an ever-increasing vasodilatation of the blood vessels supplying the placenta throughout the course of gestation. However, at any given moment during the second and third trimesters, essentially no further acute vasodilatation of these vessels can occur. Therefore, to maintain constant oxygen delivery during episodes of reduced PBF, increased oxygen extraction must occur (Fig. 3). Based on reported data, it would appear that the limit of increased extraction is twofold. Thus, PBF has to be reduced more than 50% before oxygen delivery is diminished. These considerations assume that the rate of homeostatic PBF was optimal prior to the flow reduction. We know, however, that homeostatic levels may be suboptimal either sub-acutely, as in toxemia, or chronically, as in nephropathies. In such cases, the safety factor, that is, the percent that PBF can be reduced before oxygen delivery is compromised, is lowered. This must always be considered when one evaluates the effect of an acute stimulus on the fetus.
|
*In man and subhuman primates, the spiral arterioles that will supply the intervillous space (approximately one per placental cotyledon) lose their capillary connections and dilate about tenfold as the result of replacement of the vessel walls by trophoblast. This process may extend to or below the myoendometrial junction, literally turning the spiral arterioles into spiral arteries. Blood flowing through these modified blood vessels into the intervillous space comprises PBF.
In the ovine species, multiple discrete areas of endometrium called caruncles are present in the nonpregnant state. During pregnancy, interdigitation of fetal and maternal tissues occurs in these caruncles, which enlarge to become individual placental cotyledons. Blood flowing to these areas collectively comprises PBF. Although the cotyledonary arterial blood vessels dilate significantly, they contain no trophoblastic elements, maternal-fetal transfer is by a capillary-capillary interface, and no intervillous space is present.
Dynamic Responses of the Uterine Circulation
The blood vessels of the nonpregnant uterus respond similarly to those of any other muscular organ with the exception of their unique reactivity to estrogenic stimulation, their response to local anesthetic agents, and, possibly, their response to prostaglandins (Table 1). During pregnancy, the blood vessels supplying the placenta progressively dilate, achieving a state in which minimal or no further acute dilatation can occur.24 Therefore, stimuli which evoke vasodilatation in the nonpregnant uterus are ineffective after definitive placentation has occurred. However, the placental vessels retain their ability to vasoconstrict as in the nonpregnant state. Since PBF approximates 80% to 90% of total UBF at term pregnancy, total uterine vascular responses will appear as those of the placental vessels. This point is often misinterpreted, and one must remember that the responses of the nonplacental vessels are the same whether pregnancy is present or not.
TABLE 1. Responses of the Nonpregnant Uterine Vasculature*
Vasoconstriction | Vasodilatation |
α-Adrenergic stimulation | β-Adrenergic stimulation |
(dopamine, epinephrine, | |
Acetylcholine and parasympathomimetic agents23 | |
Sympathomimetic agents11 | Adenosines17 |
(vasopressor drugs) | |
Hypoxemia(severe)12 | Cyanide19 |
Local anesthetic agents | Estrogenic substances |
(delayed but prolonged effect)2 | |
Glucosamine | |
(α-adrenergic mediation)15 | (? osmotic effect)21 |
| Hypoxemia (mild)12 |
| Ischemia19 |
| Nitroglycerine19 |
| |
| Vasoactive intestinal polypeptide20 |
* The marked dilatation of the blood vessels supplying the placenta and the preponderant distribution of uterine blood flow to the placenta obliterate gravid responses; therefore the effects of vasodilator stimuli during pregnancy are controversial. When tested, nonplacental vascular responses are similar to nonpregnant responses.
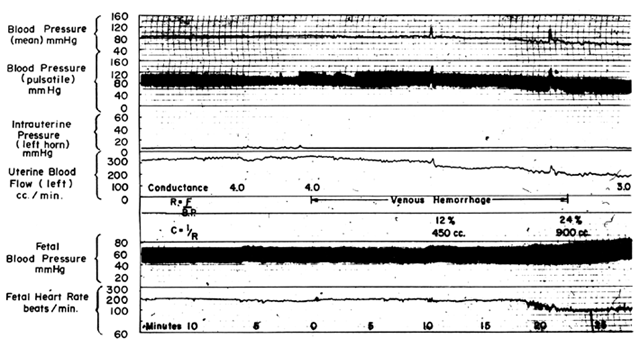
The hemochorial structure of the human placenta adds a unique factor to those that normally control blood flow in other vascular beds. That is, blood traverses the spiral arteries to enter the swamplike intervillous space, perfuses the fetal villi, and then returns to the general circulation by many collecting veins in the basal plate. Effectively, blood leaves normal vascular channels to circulate in a new extravascular space grafted onto the uterus for the duration of pregnancy. Since the intervillous space lies within the uterine cavity and since the placenta is a flexible structure, pressure generated by the contracting myometrium will be transmitted equally to the amniotic cavity and the intervillous space. Thus, a factor extraneous to usual vascular control, myometrial activity, can change intervillous space pressure and influence PBF by its effects on perfusion pressure. A schematic representation of blood flow to the non-placental tissues and to a single cotyledon is shown in Figure 4, along with the formulas pertinent to the control of each. It should be noted that these formulas are only applications of Ohm's law to these individual cardiovascular situations.
|
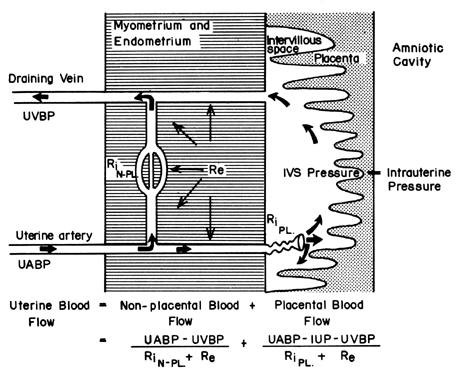
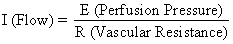
The perfusion pressure delivering blood to nonplacental tissues is the difference between uterine arterial and venous blood pressures. However, the perfusion pressure delivering blood to the intervillous space is the difference between uterine arterial blood pressure and the intervillous space pressure. The latter is best approximated by amniotic fluid pressure (IUP). In a muscular organ, resistance factors will include resistance from reactivity of vascular smooth muscle, or intrinsic resistance (Ri), and the squeeze imparted to blood vessels as they traverse the contracting myometrium, or extrinsic resistance (Re). It should be evident then that uterine contractions can affect PBF by two mechanisms: by increasing Re and by reducing placental perfusion pressure.
In clinical practice, three major characteristics of placental vascular control are important. These include the relationship between perfusion pressure and flow, the responses of the spiral arteries to vasoactive stimuli, and the effects of myometrial contractions. In addition, the unique effects of local anesthetic agents must be appreciated.
Pressure-Flow Relationship
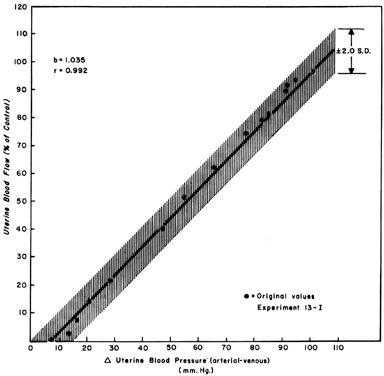
When one observes changes in UBF secondary to reductions in perfusion pressure during myome-trial quiescence, a straight-line relationship with a slope of one can be developed (Fig. 5). This reflects the widely dilated nature of the placental vasculature and indicates that PBF will decrease almost in identical proportion to the decrease in perfusion pressure. Since uterine venous pressure is quite constant under most circumstances, changes in systemic blood pressure (MBP) may be used to approximate PBF changes. That is, a 25% decrease in mean arterial pressure would be expected to cause a 25% decrease in PBF. Such measurements should be made in the lateral decubitus position, however, since pressure of the gravid uterus on the aorta alone has been shown to decrease blood pressure in the pelvic area below that observed in the brachial artery.
|
Responses to Vasoactive Stimuli
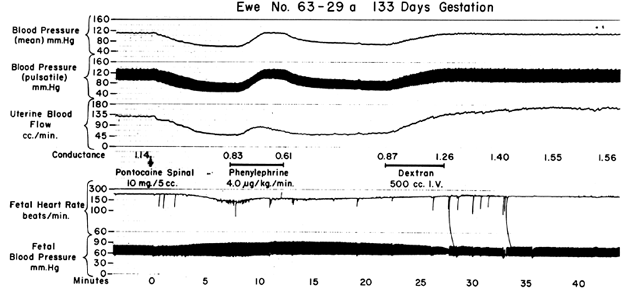
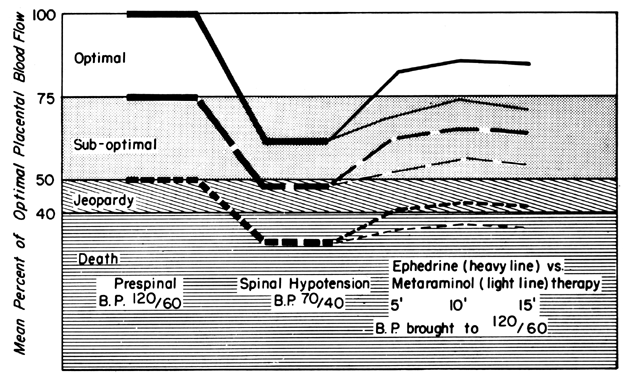
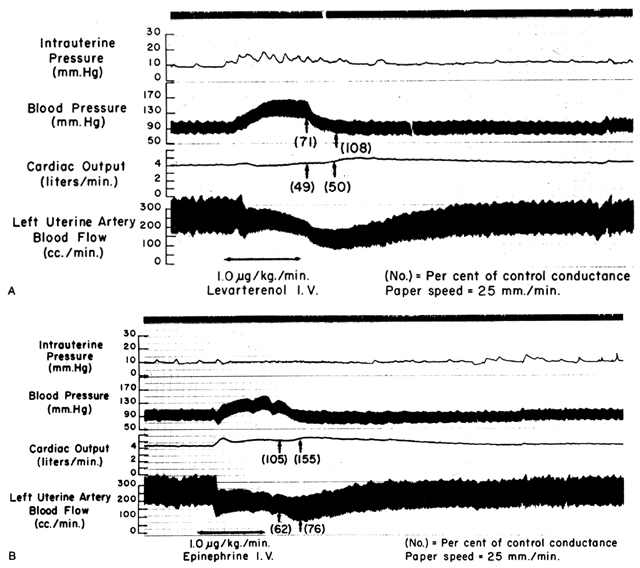
Since the spiral arteries approach maximum dilatation in the resting state, vasodilator agents or stimuli have little or no effect even though receptors for such agents are present. However, the smooth muscles of these vessels are exquisitely sensitive to vasoconstrictor agents or stimuli, more so than most other peripheral vascular beds (Fig. 6). This means that although MPB may increase in response to stimulation by a peripherally acting vasopressor drug such as phenylephrine, the proportionate increase in placental vascular resistance (Ri) is SO much greater that the net effect is a marked decrease in PBF. Such differences in vasoconstrictor sensitivity must be considered whenever a vasopressor drug is indicated. Use of a more centrally acting drug such as ephedrine, although causing a small amount of placental vasoconstriction, will result in a proportionately greater improvement in MBP with an absolute increase in PBF.
|
Myometrial Contractions
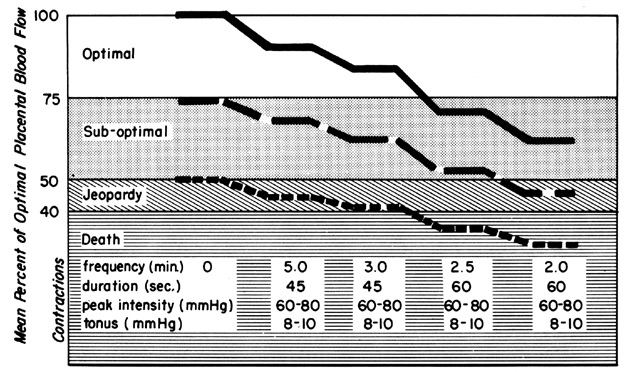
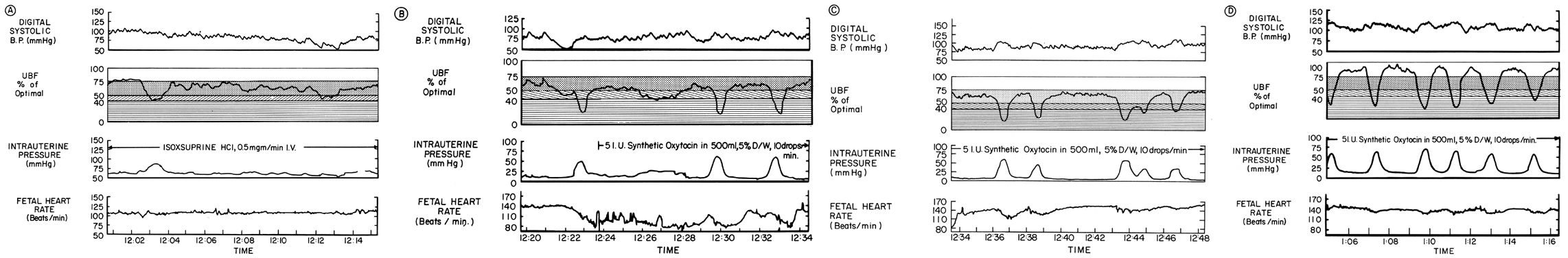
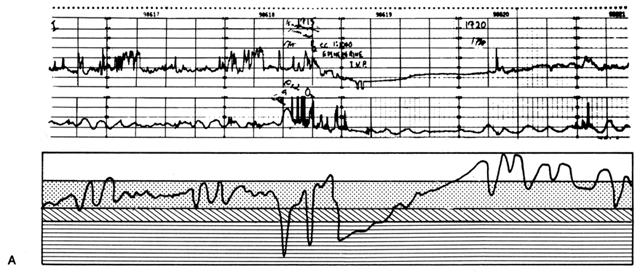
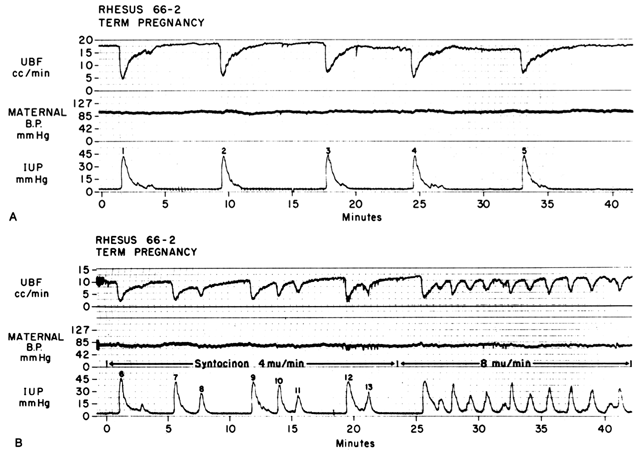
Acting by the two mechanisms discussed above, increasing Re and decreasing perfusion pressure, myometrial contractions decrease PBF in direct proportion to the intensity and duration of each contraction. The relationship is so precise that a tracing of intrauterine pressure is almost an exact inverse image of PBF (Fig. 7). Increasing the frequency of contractions decreases PBF during a given unit of time by decreasing the duration of myometrial diastole, that time when PBF is at homeostatic levels. In addition, should intercontraction tonus be elevated, as in a placental abruption, intercontraction PBF will be proportionately reduced. Radioangiographic studies in subhuman primates and women show that during the acme of myometrial contractions of average intensity, PBF ceases.25 It is evident then that labor is inherently stressful to the fetus, since the mean amount of PBF perfusing the fetal villi per given period of time progressively decreases as the frequency and intensity of uterine contractions increase.
|
Local Anesthetic Agents
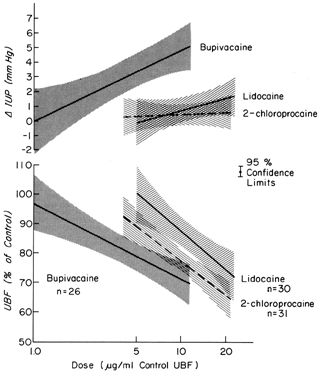
These drugs may exert effects on a vascular bed directly, as after inadvertent intravascular injection, and indirectly, as a result of paralysis of autonomic nerves that maintain normal vascular tonus. In most organs, intravascular injections have no significant effects on vascular resistance. However, the uterine and placental vasculatures respond to such stimuli with significant vasoconstriction. In addition, the myometrium is variably stimulated by such drugs (Fig. 8). Alone or together, these responses diminish PBF. Following paracervical block anesthesia administered during labor, a delayed fetal bradycardia may occur. The best hypothesis to explain the fetal response is that local anesthetic agents are injected close to the uterine arteries and that because of their excellent penetrance, they cross the arterial walls to cause their uterine effects decreasing PBF and causing fetal hypoxia.
|