Physiology and Tests of Thyroid Function
Authors
INTRODUCTION
In the embryo, the thyroid gland is derived from endodermal tissue at the base of the tongue. Thyroid tissue migrates to the anterior trachea, at the level of the cricoid cartilage, and forms a bilobed structure weighing 15–20 g in the adult. The development of the thyroid gland is under the control of at least two thyroid-specific transcription factors (TTF1 and TTF2) that have been described. Thyroid tissue remnants can be found anywhere along the path of migration. This ectopic thyroid tissue is usually hypofunctioning and is one of the causes of congenital hypothyroidism.1
The thyroid gland receives a rich blood supply via (1) arteries arising from the external carotid arteries and the subclavian arteries, and (2) innervation from the sympathetic nervous system. The recurrent laryngeal nerve lies in grooves between the trachea and the lateral lobes of the thyroid gland.
The functional unit of the thyroid gland, for hormone synthesis and storage, is the thyroid follicle.1 The thyroid follicle is a spherical structure with follicular cells forming the outer shell. Thyroglobulin is synthesized by these thyroid cells and secreted across the apical membrane into the follicular lumen (Fig. 1). The thyroglobulin-containing material that fills the follicle, termed colloid, serves as a storage form of iodine. Colloid is the substrate for the reactions that result in the synthesis of thyroid hormone. The activity of the thyroid is directly regulated by the action of thyrotropin (thyroid-stimulating hormone [TSH]) secreted by the anterior pituitary. TSH stimulates resorption of colloid and, under chronic stimulation, produces follicular hyperplasia and reduced follicular lumen size. Defects in thyroid hormone synthesis can result in increased colloid stores and goiter.
|
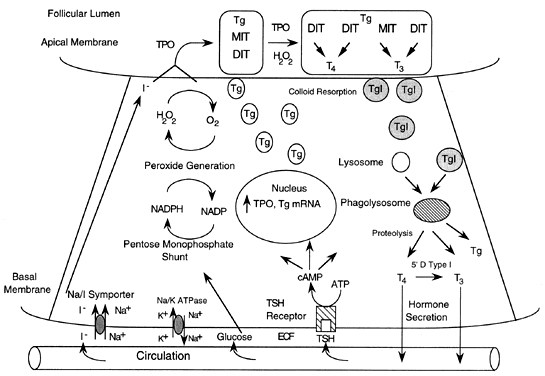
THYROID PHYSIOLOGY
Thyroid hormone influences a wide range of processes, including growth, development, maturation of the nervous system, reproduction, metabolism, and muscle function.2 The primary target organs include the brain, liver, skeletal muscle, heart, and bone. The thyroidal secretion consists of thyroxine (T4) and triiodothyronine (T3). T3 is the active form of thyroid hormone and combines with a specific nuclear receptor that binds to the regulatory region of genes and modifies expression.3, 4 Thyroid hormone synthesis and secretion is regulated by TSH secreted from the anterior pituitary, which in turn is stimulated by thyrotropin-releasing hormone (TRH) from the hypothalamus. Both TSH and TRH are regulated in a negative feedback loop by circulating T4 and T3.
Iodine, a trace element, and selenium (see later discussion) are essential for normal thyroid hormone metabolism.5 The mechanisms that regulate thyroid hormone metabolism allow for continuous production of thyroid hormone despite variation in the supply of dietary iodine. There are also maternal and fetal modifications of thyroid hormone metabolism associated with various stages of pregnancy that maintain a euthyroid state for mother and fetus.6
Iodine Transport
The majority, 70–80%, of the 15–20 mg of total body iodine is contained in the thyroid.5 To produce the daily requirement of thyroid hormone, the thyroid must trap approximately 60 mg of iodine from the circulation each day. The iodine that is not trapped by the thyroid is excreted in the urine and closely matches dietary intake. Iodine uptake in the thyroid is an active process, driven by the Na+ gradient generated by Na+/K+ adenosine triphosphatase (see Fig. 1). The Na+/I- symporter is a membrane-bound protein located in the basolateral portion of the thyroid follicular cell. The symporter passively transports two Na+ and one I- down the Na+ ion gradient, producing an iodine gradient from the thyroid cell to the extracellular fluid of 100:1. In conditions of iodine deficiency, a gradient as high as 400:1 can be generated. Other tissues that concentrate iodine include the salivary gland, gastric mucosa, lactating mammary gland, ciliary body of the eye, skin, placenta, and choroid plexus. Iodine transport is stimulated by TSH via cyclic adenosine monophosphate (cAMP) and can be competitively inhibited. Inhibitors of iodine transport include natural dietary 'goitrogens', such as the cyanogenic glucosides found in cassava, a staple in many parts of Asia and Africa, as well as perchlorate and high concentrations of iodine.
The primary source of iodine is in the diet, about 250 mg daily in the United States, but there are a number of areas worldwide, especially mountainous regions of Africa, Asia, and some parts of Europe, with dietary iodine deficiency.7 The offspring of iodine-deficient woman have a high incidence of cretinism with severe neurologic deficit and growth retardation. Even in areas of marginal iodine insufficiency, such as Belgium, there is a reduction in maternal circulating thyroid hormones and an elevation in serum TSH concentration compared to pregnant women from iodine-sufficient areas.6 The influence of these changes on fetal development are unknown.
Thyroid Hormone Synthesis
Thyroid hormone synthesis requires a number of components, including iodide, thyroid peroxidase, thyroglobulin, and hydrogen peroxide (H2O2).1 Iodine is transported into the thyroid in the inorganic form, is oxidized by the thyroid peroxidase-H2O2 system and then used to iodinate tyrosyl residues in thyroglobulin (see Fig. 1). T4 and T3 are produced by coupling of iodinated tyrosyl intermediates, which are then hydrolyzed and secreted into the circulation. The function of this system requires that these processes are closely linked, and defects in any of the components can lead to impairment of thyroid hormone production or secretion.
Thyroglobulin, a large dimeric glycoprotein (660 kilodaltons [kd]), is the major iodine-containing protein of the thyroid.8 It serves as a storage form of accessible thyroid hormone for the circulation. A small amount of thyroglobulin can be detected in the serum and is, in general, proportional to the thyroid mass. The generation of H2O2 is essential for the iodination of thyroglobulin and subsequent coupling reactions. Reduction of molecular oxygen to H2O2 requires reduced adenosine nucleotide, nicotinamide adenine dinucleotide phosphate (NADPH), and NADPH oxidase. Thyroglobulin serves as a substrate for coupling of monoiodotyrosine (MIT) and diiodotyrosine (DIT) by the thyroid peroxidase-H2O2 system. A tyrosyl phenolic ring is cleaved and joined to an iodinated tyrosine by an ether linkage. Coupling of two DIT moieties forms T4, and coupling of MIT and DIT forms T3. The thyroidal secretion usually contains approximately 80% T4 and 20% T3, but this ratio can be altered by a number of conditions, including the extent of TSH stimulation and iodine status. The TSH receptor-stimulating IgG of Graves' disease increases the fraction of T3 in the thyroidal secretion by preferential MIT/DIT coupling. Additionally, intrathyroidal type I 5´-deiodinase accelerates conversion of T4 to T3 (see later discussion).
Thyroid hormone secretion begins with resorption of colloid at the apical membrane. Pinocytosis produces multiple vesicles containing colloid, which fuse with lysosomes to form phagolysosomes. The resulting proteolysis in these vesicles releases iodothyronines, which diffuse across the cell and into the cytoplasm.
Thyroid peroxidase is a membrane-bound glycoprotein that catalyzes the oxidation of iodine, iodination of tyrosine residues, and coupling of iodothyronines.9 The human thyroid peroxidase gene is 150 kilobases (kb) and is located on chromosome 2. The predicted protein consists of 933 amino acids and has transmembrane domains at the carboxy terminus. TTF1 and TTF2 regulate thyroid peroxidase gene expression. Circulating IgG antibodies to thyroid peroxidase are pathogenic in a number of thyroid diseases, including Hashimoto's disease and postpartum thyroiditis. These antibodies can damage the thyroid follicular cells directly by activating the complement cascade.
Thyroid Hormone Transport
Thyroid hormones are hydrophobic, and the vast majority of hormone circulates bound to serum proteins with only a small free fraction of T4 (0.02%) and T3 (0.30%).10 The free hormone is the only metabolically active fraction of the total hormone concentration. The predominant thyroid hormone binding serum protein is T4-binding globulin (TBG), which carries approximately 70% of serum T4 and the majority of T3. Transthyretin (previously called T4-binding prealbumin) binds approximately 20% of T4 and also transports retinol by forming a complex with retinol-binding protein. The remaining serum T4 and T3 are bound by albumin. All three of these proteins are made in the liver, and their levels—as well as levels of total thyroid hormone—are influenced by liver disease (elevated in hepatitis, reduced in chronic liver insufficiency) and renal disease (excessive loss in the urine in nephrotic syndrome). TBG consists of approximately 20% carbohydrates by weight, and the extent of sialylation influences the circulating serum half-life (range, 15 minutes to 3 days). Estrogen increases the extent of sialylation and increases serum TBG concentration by prolonging the circulating half-life.11 Mutations of the TBG gene, located on the X chromosome, can result in partial or complete deficiency. Affected persons have reduced total thyroid hormone concentration but normal levels of free hormone and are clinically euthyroid.
Thyroid Hormone Metabolism
The thyroidal secretion is primarily the prohormone T4, which is metabolically inactive and must be converted to the active hormone T3 by removal of the 5´ phenolic (outer) ring iodine by 5´-deiodinase (Fig. 2).12, 13 Approximately 20% of circulating T3 is derived from thyroidal secretion and 80% from peripheral conversion of T4, primarily in the liver, muscle, and kidney. T4 can also be converted to reverse T3 (rT3), which is metabolically inactive; however, when elevated in the serum, it is usually associated with reduced T4 to T3 conversion (e.g. in severe nonthyroidal illnesses). Degradation products (containing 3–0 iodides) undergo sulfation and glucuronide formation for excretion in urine and bile.
|
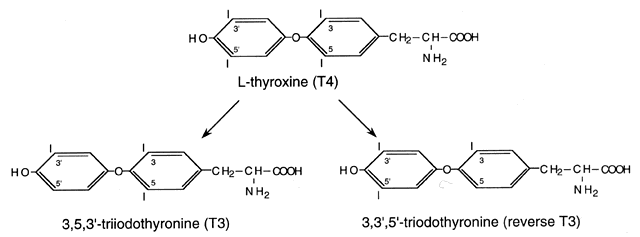
The deiodinases can be separated into outer ring (phenolic) 5´-deiodinases (types I and II) and inner ring (tyrosyl) 5-deiodinases.13 All deiodinases have a characteristic pattern of expression during development and in adult tissues, substrate preference, direction of T3 regulation, and sensitivity to inhibitors (Table 1). All deiodinases share the property of being selenoproteins that contain the amino acid selenocysteine, essential for full catalytic activity.12 The trace element selenium is required for normal thyroid hormone metabolism, and abnormal thyroid function is seen in areas of selenium deficiency, such as China and Africa.
Table 1. Properties of iodothyronine deiodinases
| Type I 5'-Deiodinase | Type II 5'-Deiodinase | Type III 5-Deiodinase |
Tissue distribution | Thyroid, liver, kidney | Brain, pituitary, brown fat, thyroid | Placenta, developing brain, skin |
Preferred substrate | rT3>>T4> T3 | T4> T3 | T3(sulfate) > T4 |
Target | Outer ring | Outer ring | Inner ring |
Response to T4 | Increase | Decrease | Increase |
Inhibition by PTU | Yes | No | No |
Inhibition by iodine contrast agents | Yes | Yes | Yes |
Physiologic role | Extracellular T3 production | Intracellular T3 production | Inactivation of T4 and T3 |
T4 = thyroxine; T3 = triiodothyronine; rT3 = reverse T3; PTU = propylthiouracil
Type I 5´-deiodinase is found predominantly in liver, kidney, skeletal muscle, and the thyroid and converts T4 to T3. It requires reduced thiol as a cofactor and is sensitive to inhibition by propylthiouracil, starvation, illness, glucocorticoids, and propranolol. The type I gene is positively regulated by T3, and its activity increases in hyperthyroidism.
Type II 5´-deiodinase is predominantly found in the pituitary, brain, brown fat, and human thyroid. The brain, unlike the periphery, derives the majority of T3 locally from T4 by the action of type II deiodinase. In hypothyroid states, type II 5´deiodinase activity increases, presumably to sustain a T3 supply to the brain. Type II 5´-deiodinase is insensitive to propylthiouracil but is inhibited by iodine-containing contrast agents, such as iopanoic acid.
Type III 5-deiodinase is found in the placenta, developing brain, and skin. It inactivates T4 and T3 by removal of a tyrosyl ring iodide. Like type I, the type III gene is positively regulated by T3. This enzyme is expressed very early in development and may be important in regulating maternal and fetal thyroid hormone metabolism.
The Hypothalamic-Pituitary Axis
The primary regulator of thyroid hormone production is TSH. TSH is a heterodimeric protein that shares a common alpha subunit with the other anterior pituitary glycoproteins (follicle-stimulating hormone, luteinizing hormone [LH], and human chorionic gonadotropin [hCG]), but has a unique beta subunit.14 The TSH alpha and beta subunit genes are both transcriptionally regulated by T3, and sequences in the 5´-flanking region have been identified that bind thyroid hormone receptor and confer these effects. Although elevated circulating T3 levels alone will suppress TSH, circulating T4, via intrapituitary type II 5´-deiodinase, is thought to be the primary regulator of TSH. T4 and T3 also provide feedback to TRH. TRH stimulates TSH as well as prolactin, and T3 inhibits expression of the TRH gene.
TSH acts via the membrane-bound TSH receptor.15 TSH is a classic seven-transmembrane structure G-protein coupled receptor. Stimulation of the TSH receptor promotes protein iodination as well as thyroid hormone synthesis and secretion. The majority of these effects are the result of enhanced intracellular cAMP accumulation. Point mutations in the TSH receptor gene have been reported that inactivate as well as those that constituitively activate the receptor, resulting in clinically significant hypothyroidism and hyperthyroidism, respectively. Acquired activating mutations of the TSH receptor gene have been reported in a high fraction of autonomously functioning thyroid adenomas. TSH is mitogenic and promotes thyroid growth, as is seen in Graves' disease and some goiters. The similarity of the TSH receptor to the LH/hCG receptor is thought to be the basis for thyroid simulation in high hCG states, such as hydatidiform mole and hyperemesis gravidarum.16
There is a log-linear relationship between serum TSH and T4 concentrations. This is an important principle to remember when adjusting T4 replacement or following thyroid function tests over time. A small change in serum T4 can produce a large change in TSH. This relationship is likely designed to achieve tight control of serum thyroid hormone levels around physiologic levels.
THYROID FUNCTION TESTS
In the last few years the development of more sensitive laboratory techniques for the evaluation of thyroid tests has improved the physician's ability to diagnose thyroid diseases in the office setting. A list of the most common and useful tests is provided in Table 2.
Table 2. Tests of thyroid function
Evaluation of the Pituitary-Thyroid Axis
Thyroid-stimulating hormone (TSH)
Circulating Levels of Thyroid Hormones
Total thyroxine (TT4)
Total triiodothyronine (TT3)
Free thyroxine (FT4)
Free triiodothyronine (FT3)
Thyroid Hormone Binding Tests
Resin T3 uptake (RT3U) or resin uptake ratio (RUR)
Indirect Measurements of Thyroid Hormones
Free thyroxine index (FT4I)
Free triiodothyronine index (FT3I)
In Vivo Isotopic Tests
Thyroid radioiodine uptake (RAIU)
Anatomic and Pathologic Diagnosis
Thyroid scintigraphy
Ultrasonography
Fine-needle aspiration biopsy (FNAB)
Ancillary Tests
Antithyroid antibodies
Anti-TPO antibodies
Antithyroglobulin antibodies
TSH receptor antibodies (TRAb)
Serum thyroglobulin (Tg)
TPO = thyroid peroxidase
Thyroid-Stimulating Hormone
The determination of serum TSH is currently the best screening test for evaluating thyroid disease.17 The first-generation TSH radioimmunoassay detected levels in the lower range of 1–2 U/ml. Although this test was once useful in the detection of primary hypothyroidism, it has been found not to be sensitive enough to separate low or suppressed TSH values (seen in hyperthyroidism) from normal values. The newer methods are the second-generation TSH immunometric assay, which has a detection limit of 0.3–0.5 U/ml; and the third-generation chemiluminometric assay, which is approximately 10-fold more sensitive at 0.05 U/ml or lower. (A value of less than 0.05 U/ml is almost always diagnostic of hyperthyroidism.) These tests offer, in addition to their greater sensitivity and specificity, the advantage of obtaining the final result in a shorter period of time, usually within 24 hours.18 The use of the second- and third-generation TSH tests has eliminated the need to perform TSH-releasing hormone stimulation tests.
The approach recommended for screening purposes in the outpatient setting is seen in Fig. 3. In hyperthyroidism as well as in subclinical hyperthyroidism, serum TSH is suppressed. In some patients on thyroid replacement therapy and in other clinical situations, serum TSH may be suppressed as well (Table 3). An elevation in serum TSH is indicative of clinical or subclinical hypothyroidism. In patients with hypothalamic or pituitary hypothyroidism, serum TSH is usually within normal limits.
|
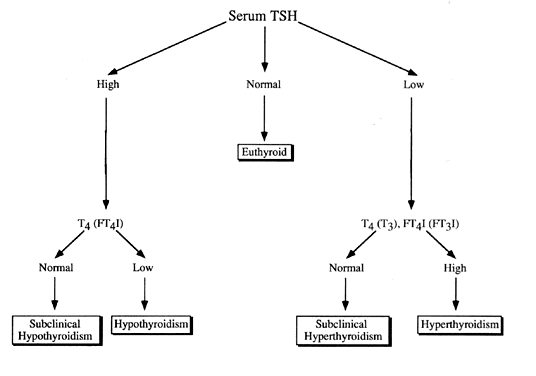
Table 3. Causes of suppressed serum TSH other than overt hyperthyroidism (normal FT4 and FT3)
Subclinical Hyperthyroidism
Treatment with thyroid hormones
Early manifestation of hyperthyroidism
Autonomous thyroid nodule
Pregnancy
Up to 15% of normal pregnancy
Multiple gestation
Nausea/vomiting mild
Transient hyperthyroidism of hyperemesis gravidarum
Hydatidiform mole
Nonthyroidal Illness
FT4 = free thyroxine; FT3 = free triiodothyronine
Total Thyroxine
Total serum thyroxine (TT4) is the most widely available test of thyroid function; however it has been replaced by the determination of serum TSH as the best practical screening test for thyroid disease. TT4 is measured by a radioimmunoassay or by fluorescence polarization assays. It detects both the T4 bound to serum proteins and the minimum amount (0.03%) that is free and biologically active. The normal range of serum TT4 is between 5 and 12.5 μg/dL, with variations in normal values according to the method used. There are no significant changes associated with diurnal variations or with differences in age or gender. Serum TT4 concentrations are elevated in almost every patient with thyrotoxicosis; however, they are also elevated in other conditions, such as alterations in TBG levels (Table 4), the presence of abnormal proteins, drugs, nonthyroidal illnesses, or the syndrome of peripheral resistance to thyroid hormones (Table 5). Therefore, TT4 determination by itself is not diagnostic of thyroid dysfunction; a better assessment of the biological effect of serum T4 is obtained by the determination of free T4 (FT4) levels, by calculation (using a determination of TBG), or by direct measurement.
Table 4. Alterations in thyroxine-binding globulin concentration
Increase | Decrease |
Pregnancy | Androgens |
Estrogen therapy | Anabolic steroids |
Acute intermittent porphyria | Acromegaly |
Infectious hepatitis | Nephrotic syndrome |
Genetically determined | Genetically determined |
Long-term perphenazine administration | |
Table 5. Causes of euthyroid hyperthyroxinemia
Binding Abnormalities
Excessive thyroxine-binding globulin (see Table 4)
Abnormal thyroxine binding to albumin (FDH)
Prealbumin-associated hyperthyroxinemia
Thyroid Hormone Resistance Syndromes
Drugs
Amiodarone
Iodinated contrast agents
Amphetamines
NonThyroidal Illness
Idiopathic
FDH = familial dysalbuminemic hyperthyroxinemia
Total Triiodothyronine
Normal values are between 70 and 180 ng/dL. As in the case of TT4, the determination of free triiodothyronine (FT3) or its calculation (see later discussion) is needed to assess thyroid status.
Free Thyroxine Index and Free Triiodothyronine Index
As mentioned before, serum TT4 and TT3 concentrations may be elevated or decreased in euthyroid persons. Therefore, an estimation of the free hormone concentration, which actually represents the free or biologically active hormone, is done by indirectly assessing the concentration of TBG in serum. In most clinical situations, the results have correlated well with the actual determination of free hormone(s) in serum.19 Exceptions are those cases with acute illness and extreme abnormalities in binding proteins. For the estimation of the free thyroxine index (FT4I), the resin T3 uptake test (RT3U) is used. RT3U is not a thyroid test per se (it is sometimes confused with the TT3 test); in conjunction with TT4 and TT3, a calculation of the amount of serum free hormone concentration is obtained. The test is carried out by adding radiolabeled T3 tracer to the patient's serum. The labeled T3 binds to available binding sites in the TBG molecules. An exchange resin or talc is added to the sample, and the labeled T3 not bound to TBG is absorbed into the anion exchange resin or talc. The result is reported as the percent of tracer bound to the resin or as an uptake ratio (sample measured resin uptake/control serum uptake) (Fig. 4).
|
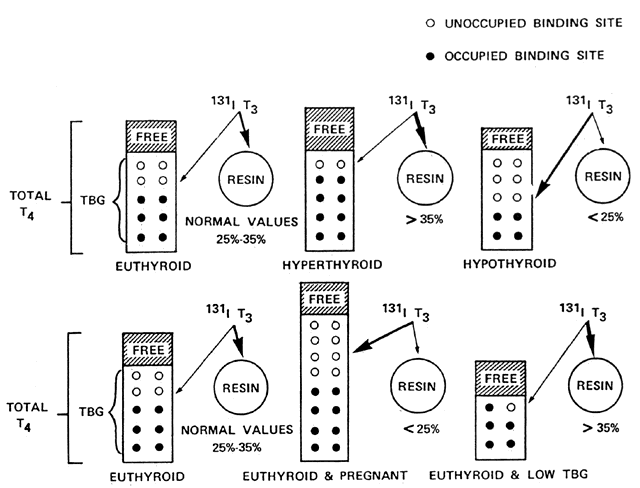
Free Thyroxine and Free Triiodothyronine
The gold standard for these tests is the equilibrium dialysis method.20 This assay is usually available only in research laboratories; it is technically difficult to perform, time-consuming, and costly. Direct-measurement methods are available in different commercial laboratories; the 'two-step' radioimmunoassay method is being used more frequently and appears to give accurate results.21 No commercial method is available, however, that accurately detects free hormone levels in patients with binding-protein abnormalities. The physician should become familiar with the methods and normal values reported by their institutions in interpreting the tests.
In Vivo Isotopic Tests
Thyroidal Radioiodine Uptake
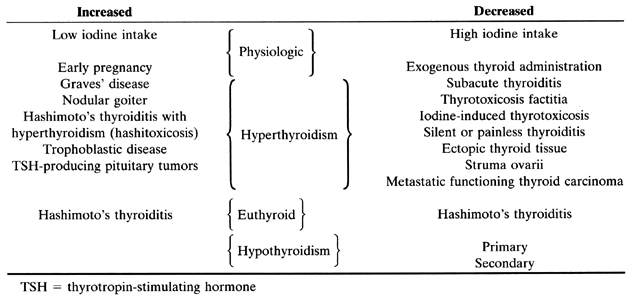
This test does not reflect the patient's thyroid status, but determines the turnover of iodine in the gland. It measures 6 or 24 hours uptake of 131I or 123I after the oral administration of the iodine. Its main use is in the differential diagnosis of the etiology of hyperthyroidism (Table 7). It should be used only when the cause of hyperthyroidism is in doubt, such as in patients with hyperthyroid symptoms, small goiters and no evidence of exophthalmopathy. It is also indicated in the hyperthyroid phase in postpartum thyroiditis to differentiate Graves' disease from destructive thyroiditis.22
Table 7. Results of 24-hour thyroid uptake in physiologic and pathologic situations
Anatomic and Pathologic Diagnosis
Thyroid Scintigraphy
Thyroid scans (e.g. using the radioisotopes 131I, 123I, and 99mTc) are not as commonly used now as they once were because of the availability of more sensitive tests for the evaluation of thyroid masses.23 Fine-needle aspiration biopsy for diagnostic cytology is usually employed as the first line of testing in the evaluation of single thyroid nodules. Thyroid scanning is used in the presence of a suppressed serum-sensitive TSH and a palpable thyroid nodule to rule out autonomous nodules. The most common use for scintigraphy is in the management of thyroid cancer after ablation therapy to detect residual or metastatic disease.24
Ultrasonography
Since the introduction of fine-needle aspiration biopsy, the role of ultrasonography in the evaluation of thyroid nodular disease has become very limited; it is also not cost-effective. It may be useful in assessing a reduction in the size of a nodular lesion during thyroid suppression therapy. Ultrasonography, as well as computed tomography or magnetic resonance imaging, of the neck area for other diagnostic purposes may detect small thyroid nodules not readily palpable because of their size of less than 1 cm. These nodules are known as thyroid incidentalomas, and in the majority of cases they need no further evaluation because most of them are benign lesions.25
Fine-needle Aspiration Biopsy
Fine-needle aspiration biopsy is a relatively simple and inexpensive diagnostic method for evaluating thyroid nodular disease.26 It is an office procedure in which a 23–25 gauge needle, sometimes with the addition of skin anesthesia, is employed. In the hands of an experienced cytopathologist, the correct diagnosis is obtained in more than 80% of cases, with an incidence of false-positive or false-negative results of less than 5%.27
Ancillary Tests
Antithyroid Antibodies
Microsomal and thyroglobulin antibodies are useful tests for the diagnosis of autoimmune thyroid disease, particularly Hashimoto's thyroiditis or chronic thyroiditis. The microsomal antigen is identical to thyroid peroxidase (TPO), a membrane-bound glycoprotein, that has been found in the luminal portion of the thyroid cell. Both antibodies—microsomal and thyroglobulin—are measured by hemagglutination or radioimmunoassay methods. These titers are positive in most persons with Hashimoto's thyroiditis: thyroglobulin antibodies in 60% of patients and antimicrosomal antibodies in 95%;28 therefore, there is no clinical indication for testing and thyroglobulin antibodies.29 Titers are elevated as well in subjects with Graves' disease. A slight increase in titer has been reported in patients after an episode of subacute thyroiditis or in patients with silent thyroiditis. In pregnant women, the presence of anti-TPO antibodies predicts the development of postpartum thyroiditis, particularly when high titers are detected in the first half of pregnancy.22
Thyrotropin Receptor Antibodies
Antibodies to the TSH receptor antibodies are immunoglobulins of the IgG family (thyroid-stimulating immunoglobulins). Most of them stimulate the thyroid TSH receptor and are present in most patients with Graves' disease. Antibodies blocking the action of thyrotropin (thyroid-blocking antibodies) have been detected in some patients with chronic thyroiditis, mainly those with atrophic autoimmune thyroiditis.30 There are several techniques for measuring these antibodies. It has been recommended that the results be reported as TSH receptor antibodies, 'measured by the specific assay'.31 There are few clinical indications for the determination of TSH receptor antibodies. In addition to the indications in pregnancy, shown in Table 8, it may be useful in the rare circumstance of exophthalmopathy and/or pretibial myxedema in a euthyroid patient.
Table 8. Indications for TSH receptor antibody determination in pregnancy
Fetal or neonatal hyperthyroidism in previous pregnancies
Active disease, on treatment with antithyroid drugs
Euthyroid, postablation or in remission, in the presence of:
Fetal tachycardia
Intrauterine growth retardation
Incidental fetal goiter on ultrasound
TSH = thyroid-stimulating hormone
Serum Thyroglobulin
Thyroglobulin is a glycoprotein normally released by the thyroid gland in small amounts. The normal concentration is 5–50 ng/ml. Its main clinical use is in following patients after ablation therapy for follicular or papillary thyroid cancer. A value of less than 5 ng/ml in patients on thyroid suppression therapy indicates little chance of tumor recurrence.32
REFERENCES
Kogai T, Brent GA: Thyroid Hormones (T4, T3). In Conn PM, Melmed S (eds): The Scientific Basis of Endocrinology: Fundamental and Clinical Principles, p 267. Philadelphia, Humana Press, 2005 |
|
Brent GA: The molecular basis of thyroid hormone action. N Engl J Med 331: 847, 1994 |
|
Williams GR, Brent GA: Thyroid hormone response elements. In Weintraub B (ed): Molecular Endocrinology: Basic Concepts and Clinical Correlations, p 217. New York, Raven Press, 1995 |
|
Yen PM, Chin WW: New advances in understanding the molecular mechanisms of thyroid hormone action. Trends Endocrinol Metab 5: 65, 1994 |
|
Dohan O, De La Vieja A, Paroder V, Riedel C, Artani M, Reed M, Ginter C, Carrasco N: The sodium/iodide symporter (NIS): characterization, regulation, and medical significance. Endocrine Reviews 24: 48, 2003 |
|
Glinoer D: The regulation of thyroid function in pregnancy: Pathways of endocrine adaptation from physiology to pathology. Endocrine Reviews 18: 404, 1997 |
|
Boyages, SC: Clinical review 49: Iodine deficiency disorders. J Clin Endocrinol Metab 77: 587, 1993 |
|
Medeiros-Neto G, Targovnik HM, Vassart G: Defective thyroglobulin synthesis and secretion causing goiter and hypothyroidism. Endocr Rev 14: 165, 1993 |
|
McLachlan SM, Rapoport B: The molecular biology of thyroid peroxidase: Cloning, expression and role as autoantigen in autoimmune thyroid disease. Endocr Rev 13: 192, 1992 |
|
Bartalena L: Recent achievements in studies on thyroid hormone-binding proteins. Endocr Rev 11: 47, 1990 |
|
Refetoff S: Inherited thyroxine-binding globulin abnormalities in man. Endocr Rev 10: 275, 1989 |
|
Berry MJ, Larsen PR: The role of selenium in thyroid hormone action. Endocr Rev 13: 207, 1992 |
|
St Germain DL, Galton VA: The deiodinase family of selenoproteins. Thyroid 7: 655, 1997 |
|
Chin WW, Carr FE, Burnside J, Darling DS: Thyroid hormone regulation of thyrotropin gene expression. Recent Prog Horm Res 48: 393, 1993 |
|
Vassart G, Dumont JE: The thyrotropin receptor and the regulation of thyrocyte function and growth. Endocr Rev 13: 596, 1992 |
|
Yoshimura M, Hershman JM: Thyrotropic action of human chorionic gonadotropin. Thyroid 5: 425, 1995 |
|
Caldwell G, Kellett HA, Gow SM et al: A new strategy for thyroid function testing. Lancet 1: 1117, 1985 |
|
Spencer CA, Takeuchi M, Kasarosyan M: Current status and performance goals for serum thyrotropin (TSH) assays. Clin Chem 42: 140, 1996 |
|
Hamada S, Nakagawa T, Mori T et al: Re-evaluation of thyroxine binding and free thyroxine in human serum by paper electrophoresis an equilibrium dialysis. J Clin Endocrinol Metab 31: 166, 1970 |
|
Surks MI, Hupart KH, Pan C et al: Normal free thyroxine in critical nonthyroidal illness measured by ultrafiltration of undiluted serum and equilibrium dialysis. J Clin Endocrinol Metab 67: 1031, 1988 |
|
Alexander NM: Free thyroxine in serum: Label thyroxine-analogue methods fall short of their mark. Clin Chem 32: 417, 1986 |
|
Brown-Martin K, Emerson CH: Postpartum thyroid dysfunction. Clin Obstet Gynecol 40: 90, 1997 |
|
Ryo UY, Vaidya PV, Scneider AB et al: Thyroid imaging agents: A comparison of 123I and 99mTc pertechnetate. Radiology 148: 819, 1983 |
|
Wong JB, Kaplan MM, Meyer KB et al: Ablative radioactive iodine therapy for apparently localized thyroid carcinoma: A decision analytic perspective. Endocrinol Metab Clin North Am 19: 741, 1990 |
|
Tan GH, Gharib H: Thyroid incidentalomas: Management approaches to nonpalpable nodules discovered incidentally on thyroid imaging. Ann Intern Med 126: 226, 1997 |
|
Hamburger JI: Consistency of sequential needle biopsy finding for thyroid nodules: Management implications. Arch Intern Med 147: 97, 1987 |
|
Gharib H: Fine-needle aspiration biopsy of thyroid nodules: Advantages, limitations and effect. Mayo Clin Proc 69: 44, 1994 |
|
Amino N, Hagen SR, Yamada N et al: Measurement of circulating thyroid microsomal antibodies by the tanned red cell hemagglutination technique: Its usefulness in the diagnosis of autoimmune thyroid disease. Clin Endocrinol 5: 115, 1976 |
|
Pearce EN, Farwell AP, Braverman LE: Thyroiditis. N Engl J Med 348: 2646, 2003 |
|
Tamakui N, Yamada T, Takasu M et al: Disappearance of thyrotropin-blocking antibodies and spontaneous recovery from hypothyroidism in autoimmune thyroiditis. N Eng J Med 326: 513, 1992 |
|
Larsen PR, Alexander NM, Chopra IJ et al: Revised nomenclature for tests of thyroid hormones and thyroid related proteins in serum. J Clin Endocrinol Metab 60: 1089, 1987 |
|
Spencer CA, Wang CC: Thyroglobulin measurement techniques, clinical benefits, and pitfalls. Endocrinol Metab Clin North Am 24: 841, 1995 |