Physiology of Puberty
Authors
INTRODUCTION
Puberty is defined as a period of transition from childhood to adulthood that encompasses physiologic, somatic, and constitutional changes associated with further development of the internal and external genitalia and secondary sex characteristics. During this period of transition, thelarche, adrenarche, pubarche, peak height velocity (PHV), and ultimately menarche occur. The culmination of pubertal development is physical maturity with the capacity to procreate. The subject of puberty can be subdivided into the somatic or physical changes associated with the external appearance of an adult and the psychological maturational process, both of which are heralded by an efflux of hormones from the adrenal glands and ovaries in girls and from the testes in boys. The culmination of these effects produces the somatic changes indicative of puberty.
SOMATIC CHANGES
Physical changes accompanying puberty have been addressed by Tanner and associates and are outlined in Tables 1 and 2 and Figure 1.1
Table 1. Breast development
Stage 1 (B1) | Preadolescent: elevation of papilla only |
Stage 2 (B2) | Breast bud stage: elevation of breast and papilla as small mound enlargement of areolar diameter |
Stage 3 (B3) | Further enlargement and elevation of breasts and areola, with no separation of their contours |
Stage 4 (B4) | Projection of areola and papilla to form a secondary mound above the level of the breast |
Stage 5 (B5) | Mature stage: projection of papilla only, owing to recession of the areola to the general contour of the breast |
Modified from Wheeler MD: Physical changes of puberty. Endocrinol Metab Clin N Am 20:1, 1991.
Table 2. Pubic hair development
Stage 1 (PH1) | Preadolescent: The vellus over the pubes is not developed further than that over the abdominal wall (i.e. no pubic hair) |
Stage 2 (PH2) | Sparse growth of long, slightly pigmented downy hair, straight or only slightly curled, appears along the labia |
Stage 3 (PH3) | Hair is considerably darker, coarser, and more curled. Hair spreads sparsely over the junction of the pubes. It is at this stage that pubic hair is first identified in the usual black-and-white photograph of the entire body; special arrangements are necessary to photograph stage 2 hair |
Stage 4 (PH4) | Hair now resembles the adult type, but the area covered by it is still considerably smaller than in the adult. There is no spread to the medial surface of the thighs |
Stage 5 (PH5) | Adult: Hair is of adult quality in terms of quantity and type, with distribution of the horizontal (or classically “feminine”) pattern. Hair spreads to medial surface of thighs, but generally not up linear alba or elsewhere above the base of the inverse triangle (overall, 10% of women have pubic hair along the linear alba) |
Modified from Wheeler MD: Physical changes of puberty. Endocrinol Metab Clin N Am 20:1, 1991.
Fig. 1. A. Pubertal development of female pubic hair. Stage 1, no pubic hair. Stage 2, sparse growth of long, slightly pigmented, downy hair, straight or only slightly curled, primarily along the labia. Stage 3, hair is considerably darker, coarser, and more curled and spreads sparsely over the junction of the pubes. Stage 4, hair, now adult in type, covers a smaller area than in the adult and does not extend onto the thighs. Stage 5, hair is adult in quantity and type, with extension onto the thighs. B. Pubertal development of female breasts. Stage 1, preadolescent. There is elevation on the papilla only. Stage 2, breast bud stage. A small mound is formed by the elevation of the breast and papilla, and the areolar diameter enlarges. Stage 3, further enlargement of breasts and areola with no separation of their contours. Stage 4, projection of the areola and papilla to form a secondary mound above the level of the breast. Stage 5, breasts resemble those of a mature woman as the areola has recessed to the general contour of the breast. (Brook CGD, Stanhope R: Normal puberty: Physical characteristics and endocrinology. In Brook CGD [ed]: Clinical Paediatric Endocrinology, p 172. 2nd ed. Oxford, Blackwell Scientific Publications, 1989.) |
Thelarche
Thelarche is the most common primary heralding sign of pubertal development. The breast bud appears at an average age of 9 years (range, 8–13 years). Asymmetry in breast development is common and best left to observation until complete breast development occurs (usually by 16–18 years of age). One variation in onset of puberty is the development of pubarche (pubic hair development) before thelarche. Unilateral onset of breast development is a recognized entity, and it may take 6 months for the opposite breast bud to become palpable.2 Some variation in the sequence of events is acceptable and perhaps normal; however, one must consider the possibility of developmental asymmetry, which may be indicative of breast tissue estrogen insensitivity or prior trauma.3
Adrenarche
At the time of birth, the androgen-producing fetal zone of the adrenal cortex undergoes involution and remains quiescent until approximately 6 years of age, when adrenarche begins. Adrenarche is defined essentially as the “pubertal transition of the adrenal gland” and is characterized by an increase in serum levels of dehydroepiandrosterone sulfate (DHEA-S), which is a direct response to adrenocorticotropin hormone (ACTH) stimulation.
Axillary Hair
On average, the presence of hair in the axilla occurs at age 13.1 years in American girls4 and 12.5 years in British girls.5 Overall, sebaceous secretion may appear at a mean age of 13.2 years.4 Further development of the apocrine glands of the axilla and vulva becomes a reflection of the increased adrenal androgens at this point.
Peak Height Velocity and Growth
PHV has been calculated by measurements of standing height taken every 3 months during the pubertal transition. PHV is the greatest increment over a period of 12 months, and the age at PHV is the midpoint of that interval. In general, PHV immediately precedes onset of menarche, with an age range of 10–15.25 years. (The mean age of PHV is 11.9 years in girls and 14.2 years in boys.6) To calculate PHV, mathematical curves are generated. Chronologic sequencing of female pubertal events is summarized in Figure 2.6, 7
Fig. 2. Sequence of pubertal events in the average American girl. (Tanner JM: Growth and endocrinology of the adolescent. In Gardner LI [ed]: Endocrine and Genetic Diseases of Childhood and Adolescents, p 14. 2nd ed. Philadelphia, WB Saunders, 1975.) |
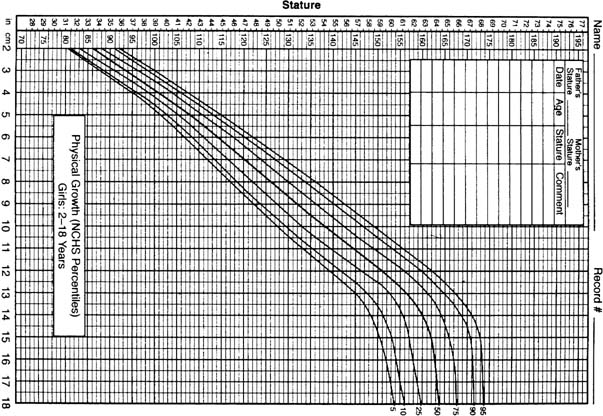
Figure 3 shows a growth chart that is used to monitor development. The pubertal growth spurt seems to be the product of multiple factors, well coordinated to achieve the normal final adult stature. These factors include estrogens, androgens, thyroid hormones, human growth hormone (hGH) (with insulin), and insulin-like growth factors (IGFs), along with their associated binding proteins working in concert with the genetic and nutritional environments. The growth spurt is integrally related to stage of sexual maturation, which normally occurs during Tanner stage 2 in girls and during Tanner stages 3–4 in boys.1, 8
{kind=link}
With the advent of recombinant hGH and its clinical use in the management of short stature (often accompanied by delayed puberty), the role of hGH in pubertal growth and development remains controversial. hGH receptors are detectable at 200 days of life, but hGH secretion begins to increase during mid childhood.9 It is secreted in a pulsatile manner and is otherwise controlled by hGH-releasing hormone (stimulatory) and somatostatin (inhibitory).9 Serum levels of hGH rise during puberty in response to increasing levels of sex steroids and their positive influence on hGH pulse amplitude.10, 11, 12 The rise in hGH pulse amplitude occurs earlier in girls than in boys, corresponding to the sex differences in puberty and growth spurt timing.13, 14 Eventually, serum levels that are two to three times baseline are achieved during puberty.9 Subsequent to these events, hGH promotes puberty by acting through IGF-I production, synergistically with follicle-stimulating hormone (FSH) at the level of the ovary to produce enhanced output of estrogens and facilitate follicle development during the menstrual cycle.13, 15
It is likely that hGH and sex steroids are necessary to achieve an optimal growth spurt; however, patients achieve greater height with sex steroids alone than with exclusive administration or production of hGH.16, 17 This interrelated responsibility in achieving a pubertal growth spurt has been concluded by observations of the stature of patients with hGH deficiency, Turner’s syndrome, and delayed or precocious puberty, along with their responses to hGH supplementation, gonadotropin-releasing hormone (GnRH) agonists, and supplemental sex steroids.9, 10, 18, 19
It has been difficult to identify a one-to-one relationship between the hGH level changes of puberty and PHV.10 Many simultaneous influences are present that could interfere with this phenomenon, including hGH-binding protein, IGF-I, and IGF-binding proteins. hGH binds to its receptors and stimulates granulosa cells to produce IGF-I, which increases during puberty and mediates many of the effects of hGH, including the augmentation of FSH at the ovarian level. IGF-I levels also may rise during puberty secondary to production via other tissues in the body, including the liver.20 Maximum IGF-I levels coincide with PHV, and it may be that these growth factors, along with their binding proteins, have direct actions on the epiphyseal growth plate and cartilage, as do estrogens.20, 21 IGF-I function is modulated by insulin.22 Insulin levels rise during puberty along with levels of IGF-I, IGF-binding protein-3, and hGH. hGH is an insulin antagonist at postreceptor sites, and it has been suggested that the rising levels of hGH identified during puberty may lead to the normal phenomenon of adolescent insulin resistance.23, 24, 25
Insulin resistance is restricted to glucose metabolism, allowing amplified protein metabolism to occur through the higher insulin levels seen during puberty, favoring growth. Another suggested way in which insulin may contribute to pubertal growth is through its inverse relationship with IGF-binding proteins and sex hormone–binding proteins. Hyperinsulinemia yields low levels of these carriers during puberty, augmenting bioactive IGF-I and sex steroids.22
In addition to their influential role in achieving growth through increasing hGH secretion, estrogens promote the growth spurt by acting directly on epiphyseal growth plates.16, 23, 24 It is well known that high levels of estrogens can accelerate skeletal maturation and fuse growth plates, inhibiting growth and effectively reducing the final adult height.25 As girls approach puberty, estradiol levels appear to be higher than in boys, the assumption being that the increased secretion of estradiol from the ovaries results in a more rapid epiphyseal development and closure in girls compared with boys. In a study by Kline and coworkers26 in which 23 prepubertal boys and 21 prepubertal girls were evaluated, the estrogen levels in girls (0.6 ± 0.6 pg/mL) were significantly greater than in boys (0.08 ± 0.02 pg/mL). An ultrasensitive recombinant cell bioassay was used for this determination. In boys, testosterone and hGH seem to be necessary elements for the pubertal growth spurt.10, 17, 24, 27 The more profound and consistent role of estrogens versus androgens in the pubertal growth spurt and attainment of final height in females has been shown clinically through observations of patients with sex hormonal and sex chromosomal abnormalities.28
Nutrition and exercise each play a role in pubertal growth and attainment of final adult height. Patients with anorexia nervosa or malnutrition can experience stunted or arrested growth.29 The precise mechanism is unclear, but a blunted hGH response to clonidine and decreased IGF-I levels have been shown in anorexic girls.29 To complicate matters further, it may be that short stature predisposes these patients to the development of anorexia nervosa.30
Elite Athlete and Growth
Acute exercise elicits a transient rise in hGH; however, intensively trained adolescent female gymnasts often have a shorter stature than age-matched competitive swimmers. Decreased height for bone age and a decreased growth velocity have been identified.31 Estradiol and IGF-I levels may be decreased, but otherwise the cause of the stature findings is unclear. Proposed mechanisms include a somatotype bias by the sport (i.e. to excel, one should be relatively short-statured), psychological stress, hGH deficiency, growth plate trauma, changes in thyroid function, and nutritional issues.32 If the epiphyseal plate has not fused already, “catch up” growth can be experienced when training is stopped. The control of growth is summarized in Figure 4. The role of thyroid hormones is reviewed later.
Fig. 4. Control of cellular growth. (Hintz RL: Abnormalities in growth. In Sanfilippo JS, Muram D, Lee PA, Dewhurst J [eds]: Pediatric and Adolescent Gynecology, p 4. 2nd ed. Philadelphia, WB Saunders, 2001.) |
Menarche
Although on the average menarche occurs at 12.8 years of age in the United States, a range of 10.7–16.1 years has been reported.33 In general, menarche occurs 24 months after thelarche and relatively soon after PHV. Approximately 18 months is required between initial onset of menarche and regular ovulatory cycles. Of all menstrual cycles in “normal adolescents” 12–14 years old, 90% are anovulatory.3
Summary of Somatic Events
On average, thelarche occurs at 9 years of age and precedes comparable development in boys (i.e. genital development) by 1 year. PHV remains an early pubertal event in girls and a relatively late one in boys. In general, the culmination of puberty in girls is menarche, which occurs at an average age of 12.7 years (range, 10.7–16.1 years). Chronologic age and bone age seem to be well correlated with circulating estradiol levels, and the mean plasma levels are different for each pubertal stage. Although the range of normal variation is quite varied from individual to individual, there remains a significant degree of overlap corresponding to specific Tanner staging. Compared with chronologic age, the Greulich and Pyle34 bone age estimates are correlated better with pubertal changes and offer a better reflection of normal physiologic development.
PUBERTAL PHYSICAL CHANGES IN REPRODUCTIVE ORGANS
External Genitalia
The vagina increases in length before the onset of secondary sex characteristics and continues to enlarge until menarche or later.3 There are accompanying cytologic changes involving the vaginal epithelium. These changes result in an increase in superficial cells compared with parabasal and intermediate cell forms, which represents an estrogen-stimulatory effect. Vaginal pH becomes more acidic approximately 1 year before menarche, representing the presence of lactobacilli.3 Physiologic leukorrhea is frequently a hallmark indicative of the impending onset of menarche.35 The mons pubis increases in size with the development of fat deposition and further enlargement of the labia majora.3 A particularly wrinkled appearance of the labia becomes more obvious, the clitoris increases slightly in size, and the urethral hillock becomes more prominent.3
Uterus
Correlation exists between Tanner score and the uterine and ovarian volumes.36 Initially, there is a 50:50 size ratio between the uterine fundus and the cervix. This association changes during pubertal development, the cervix ultimately becoming one third the size of the entire uterus. During childhood, the uterus usually lies in the midplane position. Later stages in pubertal development primarily involve the myometrium, which is a reflection of response to ovarian hormonal stimulation. The endometrium further develops after onset of secondary sex characteristics with resultant menstruation (Table 3).
Table 3. Organ development
Infancy | Prepuberty | Postpuberty |
Uterus 2.5 mm | Volume 1.7 cm3; 3.3 × 1.3 × 0.8 cm | 60 mm long |
Ovary 1.5 × 3 × 2.5 mm* | Volume 4–6 cm3; 4.0 × 2.0 × 1.5 cm |
* 5% of neonatal ovaries have cysts 1.7 mm in diameter.
Sanfilippo JS, Lavery JP: The spectrum of ultrasound: Antenatal to adolescent years. Semin Reprod Endocrinol 6:45, 1988.
Ovaries
There seems to be a continuous growth of the ovaries throughout childhood.37 There is an increase in the prevalence of polycystic ovary syndrome (PCOS) from childhood to adolescence: In a normal patient population assessed in a retrospective study, the increase was from 6% at age 6 to 26% at age 15.37 Most women with PCOS developed the problem during childhood and puberty. hGH has a definite role in ovarian growth. There also seems to be a close association between the size and appearance of the ovaries on ultrasound and specific changes in circulating levels of estrogens (Table 4).38
Table 4. Ovarian volume and morphologic aspects of the ovaries in relation to chronologic age*
Ovarian volume (cm3) | Microcysts (<9 mm) | Macrocysts (>9 mm) | ||||||
| Chronologic age (years) | Pubertal and prepubertal girls | No. | Prepubertal girls | No. | Pubertal girls | No. | No. (%) | No. (%) |
| 2 | 0.75 (0.41) | 5 | 0.75 (0.41) | 5 | — | — | 0 (0) | 0 (0) |
| 3 | 0.66 (0.17) | 6 | 0.66 (0.17) | 6 | — | — | 0 (0) | 0 (0) |
| 4 | 0.82 (0.36) | 14 | 0.82 (0.36) | 14 | — | — | 2 (14.3) | 0 (0) |
| 5 | 0.86 (0.03) | 4 | 0.86 (0.03) | 4 | — | — | 0 (0) | 0 (0) |
| 6 | 1.19 (0.36) | 9 | 1.19 (0.36) | 9 | — | — | 1 (11.1) | 0 (0) |
| 7 | 1.26 (0.59) | 8 | 1.26 (0.59) | 8 | — | — | 2 (25) | 0 (0) |
| 8 | 1.06 (0.50) | 10 | 0.90 (0.27) | 8 | 1.68 (0.87) | 2 | 2 (20) | 0 (0) |
| 9 | 1.98 (0.76) | 11 | 2.15 (0.92) | 7 | 1.69 (0.27) | 4 | 6 (54.5) | 0 (0) |
| 10 | 2.22 (0.69) | 12 | 2.23 (0.86) | 7 | 2.20 (0.47) | 5 | 6 (50) | 0 (0) |
| 11 | 2.52 (1.30) | 12 | 2.32 (0.39) | 2 | 2.56 (1.43) | 10 | 7 (58.3) | 0 (0) |
| 12–13 | 3.95 (1.70) | 10 | — | — | 3.95 (1.70) | 10 | 6 (60) | 2 (20) |
*Values are mean (SD).
Salardi S, Orsini LF, Cacciari E, et al: Pelvic ultrasonography in premenarchal girls: Relation to puberty and sex hormone concentrations. Arch Dis Child 60:120, 1985.
Male Pubertal Development
The physical signs of puberty in boys include development of the external genitalia, increase in testicular volume, presence of pubic and axillary hair, PHV, and “breaking of the voice.” Table 5 lists stages of genital development in boys.
Table 5. Staging of genital development in boys
Stage 1 (G1) | Preadolescent: testes, scrotum, and penis are about the same size in proportion as in early childhood |
Stage 2 (G2) | Enlargement of scrotum and of testes. The skin of the scrotum reddens and changes in texture. Little or no enlargement of penis occurs at this stage |
Stage 3 (G3) | Enlargement of penis, which occurs at first mainly in length. Further growth of testes and scrotum occurs |
Stage 4 (G4) | Increased size of penis, with growth in breadth and development of glans. There is further enlargement of testes and scrotum and increased darkening of scrotal skin |
Stage 5 (G5) | Genitalia are adult in size and shape |
Marshall WA, Tanner JM: Variations in patterns of pubertal stages in girls. Arch Dis Child 44:291, 1969.
In 96% of normal boys, the initiation of pubertal development is characterized by testicular enlargement. Testicular volume is 1–3 mL in the prepubertal boy and, on the average, 3.3 mL in stage 1 of pubertal development. No further enlargement takes place after stage 5 is reached; it seems that the penis size decreases slightly from the immediately postadolescent peak.6 Early puberty in boys is comparable to the period of ovarian enlargement in girls preceding onset of thelarche, perhaps in contrast to a continued slow growth of ovaries throughout childhood.36 With respect to gonadotropins, there seems to be an increase in amplitude and frequency of luteinizing hormone (LH) during puberty. There is a distinct “sex difference” with respect to FSH and LH output during pubertal development.38
As a rule, pubic hair appears soon after onset of testicular development and is complemented by the presence of axillary and facial hair. There is normally a 2-year interval between onset of pubic hair (stage 2) and the presence of axillary and facial hair. Under normal circumstances, facial hair development occurs at a mean of 14.9 years. Breaking of the voice at the time of puberty is indicative of androgen-induced enlargement of the larynx. Completion of the voice change (deepening) is complemented by finalization of pubertal development. During this pubertal process, the capability for procreation is established with development of the seminal vesicles, the prostate gland, the bulbourethral glands, and the onset of testicular spermatogenesis. The average age of first ejaculation of semen occurs in early to mid puberty (bone age, 13.5 years, or chronologic age, 14.3 years). Chronologic sequencing of male pubertal events is summarized in Figure 5.
Fig. 5. Sequence of pubertal events in the average American boy. (Tanner JM: Growth and endocrinology of the adolescent. In Gardner LI [ed]: Endocrine and Genetic Diseases of Childhood and Adolescents, p 14. 2nd ed. Philadelphia, WB Saunders, 1975.) |
A physiologic gynecomastia may occur. It is often unilateral, but it can be symmetric. In general, it occurs during genital stages 3 and 4 and regresses within an average of 12–18 months of its initial appearance. Persistence of gynecomastia necessitates surgical intervention. Generally in boys, testosterone levels increase progressively throughout all stages of pubertal development, as shown in Figure 6. In addition, FSH increases until mid puberty and can be correlated with wolffian tubular development and spermarche (see Fig. 6).
Fig. 6. Pubertal stage. (Nottelmann ED, Susman EJ, Dorn LD, et al: Developmental processes in early adolescence. J Adolesc Health Care 8:246, 1987.) |
Testosterone is the primary sex steroid produced by the testes. It is converted peripherally, at the target organ, to dihydrotestosterone. The adult testes secrete 6 mg of testosterone daily. It is present initially in the human fetus at 12 weeks’ gestation in response to human chorionic gonadotropin. At puberty, testosterone is responsible for growth of the external genitalia in boys. The wolffian ducts respond to increased testosterone production, and external virilization is primarily a phenomenon of dihydrotestosterone, as is penile urethral prostate development. Specific androgen receptors result in increased mRNA, reflecting the transcription of structural genes. In this way, androgens have an anabolic effect. The Leydig cells of the testes are responsible for steroid production. There seems to be a specific tempo regarding pubertal events.39 The range for age of onset is 9–14 years, which usually results in complete pubertal development in 2–4.5 years (mean, 3.25 years).2, 39, 40
Psychosocial Development
Perhaps the biggest challenge facing the clinician is the correct approach in handling the psychological aspects of pubertal development—a concept that must be addressed from the perspective of the patient and the parent. Any effort to facilitate communication between the health care professional and the patient and between the patient and the parent is time well spent. Objectives should be channeled so that the clinician is not perceived in any way as impersonal, “mechanical, intimidating, or [being] a poor listener.”41 Clinicians should show appropriate and genuine interest in each adolescent patient. For adolescents, perception of identity from the moment they identify their first hyperactive sebaceous gland to the time they realize they are truly adults is an important, often all-encompassing, period in their lives. Knowledge and understanding of advances in the physiologic and psychological process of puberty provide an excellent foundation for present and future gynecologic care.41
The chronologic aspects of puberty can be highlighted by an understanding of thelarche, adrenarche, PHV, and ultimately menarche. One goal of the clinician should be to have the patient experience a positive attitude and self-image in terms of the changes associated with puberty. Specific goals are for the health care provider to be supportive of the adolescent's sexual self and to encourage an atmosphere of supportive parents and an interested and concerned physician, with the ultimate goal being a positive gender role and sexual orientation.42
The changes that occur during pubertal development can be characterized on three levels.43 The first level is characterized by enhanced secretion of gonadotropins and corticotropic hormones from the anterior pituitary and is governed by genetic tenets.44 The second level is characterized by ovarian hormone production. The third level is characterized by changes in secondary sex characteristics. Deviations from what is accepted as “normal adolescent development” may result in significant psychological effects ranging from uneasiness and self-consciousness to severe embarrassment.41 The perceptions of the obese adolescent and the underweight adolescent merit attention because both have specific psychological effects. The adolescent may become passive in social interactions, resulting in many sequelae. The health care provider must identify these problems and deal with them either on a one-on-one basis or by appropriate referral. Support and counsel of the adolescent and the adults with whom the patient interacts (e.g. parents, teachers, other significant adults) set the stage for less stressful adjustments during adolescence.
Knowledge of the various stages of pubertal development from the psychological aspect merits consideration. Croft and Asmussen41 offered the following advice to health care professionals:
- Assume an active role in establishing comfortable, friendly relationships, permitting discussion and an atmosphere of mutual trust.
- Take a proactive approach with sexual discussions.
- Establish separate discussions with parents and youth as a matter of routine.
- Recognize “teachable moments,” discussing topics timed to correspond with developmental cues (pointer for parents).
- Inform families regarding imminent physical and psychosocial developmental changes.
- Enhance communication skills of parents and adolescents.
- Decrease the number of questions asked. Inappropriate and challenging questions erode trust and place the physician in an awkward position of “cross examination.”
- Avoid the use of scientific terms and jargon whenever possible.
- Use a positive approach when discussing developmental changes and needed intervention.
- Increase knowledge of the family. How does the family interact?
- Facilitate family discussions about sexuality.
- Recognize that “the talk” regarding sexuality is a myth. Sexuality topics should be discussed incrementally over time.
- Use other professional staff and referrals. This is especially important depending on the circumstance, in which more expertise, competence, interest, and available time are necessary. Psychological objectives in puberty are independence, maturation, positive self-image, emotional stability, and goal orientation.
HYPOTHALAMIC-PITUITARY-OVARIAN AXIS
Any discussion of the physiology of puberty must incorporate knowledge and understanding of neuroendocrinology of the hypothalamic-pituitary-ovarian (HPO) axis. The prevalent concepts are depicted schematically in Figure 7. Puberty is one event in a dynamic process that begins in utero. GnRH is present, stimulating the release of gonadotropins by 10–18 weeks’ gestation.45, 46, 47, 48 Levels continue to rise, which results in increasing gonadal steroid production. In the fetus and term neonate, serum estrogen levels are noted to be approximately 5000 pg/mL at term, but this also reflects the conversion of fetal and maternal C-19 steroid precursors to estrogens by the placenta. The fetal adrenal gland also plays a role in estrogen production throughout fetal life. Closer to term, gonadotropin activity declines. The mechanism of the inhibition is unclear but may involve either central factors or sex steroid–related negative feedback. This negative feedback is, and will continue to be, a predominant mode of control on the hypothalamic-pituitary axis. A manifestation of in utero estrogen levels can be noted at the time of birth, when there may be palpable breast tissue, indicative of the end-organ effect of circulating estrogens. With the sudden loss of maternal and placental sex steroid contributions, the neonate experiences a resurgence of gonadotropins, and peaks or pulses may be identified for several years. Episodic sex steroid production and ovarian cysts may result.
|
Fig. 7. Schematic representation of the central nervous system (CNS)–hypothalamic–pituitary–ovarian axis. (Lee PA, Reiter EO, Kulin HE: Neuroendocrinology of puberty. In Sanfilippo JS, Muram D, Lee PA, Dewhurst J [eds]: Pediatric and Adolescent Gynecology, p 45. Philadelphia, WB Saunders, 1994.) |
Estradiol is the major estrogen (and sex steroid) in the female body. This “GnRH pulse generator” undergoes a modicum of suppression to a mid-childhood nadir, but with the advent of new fluorimetric assays, episodic gonadotropin secretion (FSH and LH) with a circadian rhythm has been shown during the prepubertal years.49, 50, 51 The axis previously thought to be quiescent is active, but it is suppressed during the “juvenile pause.” The mechanism of this central restraint is unknown. Various clinical clues have been reviewed and summarized by Styne52 through analysis of hamartoma patients and clinical states associated with increased intracranial pressure, central nervous system neoplasms, and precocious puberty. We now know that the prepubertal pituitary is highly sensitive to GnRH, capable of yielding the greatest magnitude of increase in gonadotropin levels in response to physiologically administered GnRH stimulation during this stage of development.49, 50 FSH particularly responds dramatically before puberty. The mechanism of GnRH pulsatility and control is unknown but has been postulated to involve catecholamines and glutamate along with dopaminergic, noradrenergic, and opioid neurons and their interaction with the hypothalamus.53 With the onset of puberty, nocturnal gonadotropin secretory episodes increase, primarily LH, and in essence reflect an increase in pulse amplitude.49, 50, 51 Properly timed pulsatile GnRH secretion must exist to effect appropriate FSH and LH secretion.54 Frequently an LH pulse is followed by an FSH pulse within 30 minutes, and secondarily gonadal steroids begin to rise with pubertal onset.49, 50 Whether through the concurrently increasing sex steroids and their negative feedback influence or through inhibin, the pubertal FSH response to GnRH is dampened over that of LH, resulting in a final LH-to-FSH ratio of 1:1 (versus 1:20 before puberty).49, 50, 51, 55
As puberty progresses, wake-time gonadotropin pulses increase, and sleep-associated pulses decrease to an eventual loss of diurnal variation with more advanced puberty. Regular daytime LH pulses are followed closely by thelarche, and the achievement of a mature LH pattern seems to parallel the growth spurt in both sexes.56 Age of onset of menarche and estradiol levels seem to be interrelated. In women with early menarche, there is a twofold greater follicular phase serum estradiol level compared with women whose menarche occurs at 13 years of age or later.57 Eventually, positive feedback mechanisms ensue, which give rise to ovarian estradiol–triggered LH surges, ovulation, and the establishment of regular menses (all discussed in more detail elsewhere in this volume). Many clinical entities have contributed to the understanding of the HPO axis as it relates to puberty and the aforementioned events:
- GnRH must be responsible for even prepubertal gonadotropin pulses because these are abolished with the down-regulation that results from GnRH agonist therapy.49, 50, 51
- The juvenile pause, or mid-childhood central restraint, must not crucially involve gonadal steroids because patients with gonadal dysgenesis or agenesis (Turner’s syndrome) experience the same phenomenon.58
- Anorexia nervosa patients and intensively trained (elite) athletes can experience delayed or arrested puberty, abnormal growth, or primary or secondary amenorrhea. They have been shown to have any combination of decreased gonadotropins (pulse frequency and amplitude), decreased sex steroids, and impaired gonadotropin response to GnRH. Theories to explain such phenomena have involved crucial weight parameters and opioid or catecholestrogen metabolism changes, none of which account for all patients.32, 59, 60, 61, 62 Catecholestrogens are less potent than either estradiol or estrone. They have both catecholinergic and estrogen effects. The latter effect is minimal.
Growth hormone likely interacts with the HPO axis and has a role in puberty and secondary sex characteristic development, but this area is fraught with controversy and confusion. Isolated hGH-deficient patients often experience delayed puberty with the anticipated short stature. The impact of hGH supplementation on puberty onset, timing, and rate of progression is a subject of great debate, however.9, 13, 15, 19, 63 There is suspicion that although hGH therapy does not trigger earlier puberty, it may augment the rate of progression. One could argue that such supplementation is not physiologic. Patients with McCune-Albright syndrome who exhibit precocious puberty often are found to have hypersecretion of hGH and normal gonadotropins.15 hGH supplementation may facilitate ovulation induction with human menopausal gonadotropin.15
The mechanism of puberty promotion likely involves the capability of hGH to amplify or augment gonadotropin action on the ovary, stimulating steroid production and plausibly regulating follicular development.15 This mechanism may be effected through IGF-I as a mediator.64 The possibility still exists, however, that hGH has a direct effect on the ovary.15
As mentioned earlier, insulin augments bioactive IGF-I by decreasing its binding proteins, and this has implications for ovarian steroidogenesis. Insulin’s role may not stop there, however. It also has the ability to control sex hormone–binding proteins in an inverse relationship. Consequently the rising insulin levels seen in adolescents (insulin resistance) yield increased free sex hormones and likely contribute to sexual maturation.22 Insulin also may stimulate ovarian growth.
In their 1992 review, Nobels and Dewailly22 summarized the above-mentioned pubertal interactions (Fig. 8), then elegantly generated a hypothesis to explain why PCOS develops in many women during their adolescent years (Fig. 9). The salient points are as follows:
- Hyperinsulinemia and elevated levels of IGF-I, noted during adolescence, cause theca-cell hyperplasia and excessive androgen production from the ovaries of predisposed females.
- These intraovarian androgens lead to follicular atresia with resultant lower estradiol levels but higher estrone levels through peripheral aromatization.
- This hormonal milieu causes a relatively exaggerated response of LH secretion to GnRH, reinforcing the theca-cell hyperplasia.
|
Fig. 8. Simplified scheme of the endocrine interactions that regulate pubertal development. GnRH, gonadotropin-releasing hormone; GH, growth hormone; IGF-I, insulin-like growth factor I; IGF-BP1, IGF binding protein 1; SHBG, sex hormone–binding globulin. (Nobels F, Dewailly DD: Puberty and polycystic ovarian syndrome: The insulin/insulin-like growth factor I hypothesis. Fertil Steril 58:655, 1992.) |
|
Fig. 9. Hypothetical mechanism of onset of polycystic ovarian syndrome during puberty. GH, growth hormone; IGF-I, insulin-like growth factor I; IGF-BP1, IGF binding protein 1; LH, luteinizing hormone; SHBG, sex hormone–binding globulin. (Nobels F, Dewailly DD: Puberty and polycystic ovarian syndrome: The insulin/insulin-like growth factor I hypothesis. Fertil Steril 58:655, 1992.) |
On ultrasound, ovaries can (but do not always) appear polycystic, with arrested follicles and stromal hyperplasia. In a related manner, other authors have suggested that an alteration in hGH secretion per se sets off the PCOS cascade.15 Regardless, many features of PCOS described earlier are noted in normal puberty. In some patients, normalization of the insulin resistance or increased hGH or IGF-I secretion (normally seen with attainment of final adult height) does not occur on schedule, however, and the woman is left with hyperandrogenism and anovulation and variable features of obesity, acanthosis nigricans, and polycystic ovaries.
Thyroid Hormones
Thyroid hormones are an integral and essential prerequisite for normal pubertal development. Evaluation of total and free thyroid hormone levels and thyroxine (T4)-binding globulin during puberty has been addressed by Dunger and coworkers65 in their evaluation of 20 girls and 19 boys, ranging in age from 10 to 15 years. Assessment was made at 6-month intervals; the results noted that free T4 levels were associated with a marked decrease from 10 years of age (15.7 pmol/L) to 12.5 years of age (13 pmol/L) before rising to prepubertal (15.9 pmol/L) levels at 15 years of age. The nadir occurred during pubertal developmental Tanner stages 3–4.
Total T4 levels, although having a similar pattern, showed a slight delay in the nadir (13 years of age, puberty stage 4). These findings were for girls in the peripubertal years. T4-binding globulin concentrations remained unchanged in girls but fell slightly in boys during later puberty.
Theoretically, thyroid hormones are essential because they potentiate the actions of IGF-I on cartilage66 and stimulate hGH synthesis by the anterior pituitary gland.67 One would assume that there is a significant change in the levels of free thyroid hormone in boys versus girls during pubertal development. It is well known that thyroid hormones play an important role with respect to IGF-I action on cartilage growth in an animal model64 and that they stimulate hGH synthesis. When one looks at the entire spectrum of a female’s life cycle, the continuum begins during the neonatal stage: Thyroid hormones are elevated (i.e. at normal adult levels) in the neonate, with a progressive decrease in levels between ages 1 and 15 years.68 The fall is less marked or absent in the 10–15 years age group, and T4 and thyroid-binding globulin levels are in the normal adult range. The question remains as to whether or not there is a change in thyrotropin-receptor function that is associated with pubertal development. This question is unanswered at present. Changes in thyroid hormone levels around puberty may reflect a change (i.e. increase) in hGH secretion during pubertal development.
There also seems to be a circannual rhythm with respect to thyroid-stimulating hormone secretion. A peak is noted in December according to Bellastella and colleagues.69 hGH, T4, and triiodothyronine (T3) levels did not show any similar rhythm. These investigators concluded that before puberty, thyroid hormones play a minimal role, if any, with regard to regulation of circannual thyroid-stimulating hormone periodicity (Figs. 10 and 11).
|
Fig. 10. Serum thyroxine (T4), free T4, free triiodothyronine (T3), and thyroxine-binding globulin (TBG) concentrations in a cohort of 19 girls and 19 boys between the ages of 9.5 and 15.5 years. The values represent means ± SEM. The y-axes represent the adult normal ranges. (Dunger DB, Perkins JA, Terence P, et al: A longitudinal study of total and free thyroid hormones and thyroxine binding globulin during normal puberty. Acta Endocrinol 123:305, 1990.) |
|
Fig. 11. Sequential measures of serum free thyroxine (FT4) and free triiodothyronine (FT3) concentrations against age in 19 girls. Individual values are plotted together with a line showing the quadratic regression of free T4 or free T3 against age (A). FT4 = 50 − 5.7 A + 0.2 A2; FT3 = 2.9 + 0.83 A − 0.04 A2. (Dunger DB, Perkins JA, Jowett TP, et al: A longitudinal study of total and free thyroid hormones and thyroxine binding globulin during normal puberty. Acta Endocrinol 123:305, 1990.) |
Onset
Not only is the mechanism of mid-childhood central restraint of the GnRH pulse generator still a mystery, but also the basis for the eventual loss of this inhibition (i.e. onset of puberty) is still elusive. Many etiologic theories and factors have been proposed, such as body fat composition; nutritional status; exercise and fitness level; stress; cortisol; endogenous opiates; sex steroids; inhibin and activin; melatonin, light, and climate; hGH, IGF-I, and insulin; thyroid hormone; prolactin; and vitamin D.
Some hypothalamic role generally is agreed on, and it seems that genetics likely is involved. Any theories also must account for the initial nocturnal predominance of the LH pulsatility. A brief discussion of some of the most notable discoveries is included in this chapter. For further details, see elsewhere70 (Figs. 12 and 13).
|
Fig. 12. Profile of relative mean levels of circulating luteinizing hormone (LH) and follicle-stimulating hormone (FSH) during fetal life, infancy, childhood, and puberty. Episodic release, first apparent before birth, persists into early infancy, becomes less frequent during childhood, then resurges to stimulate puberty. (Lee PA, Reiter EO, Kulin HE: Neuroendocrinology of puberty. In Sanfilippo JS, Muram D, Lee PA, Dewhurst J [eds]: Pediatric and Adolescent Gynecology, p 54. 2nd ed. Philadelphia, WB Saunders, 2001.) |
|
Fig. 13. Schematic mechanism for the control of the onset of puberty. Central restraint of hypothalamic function is maximal in mid childhood. Sex steroid inhibitory regulation of gonadotropin-releasing hormone (GnRH) is present in infancy, early childhood, and adulthood. Pulsatile secretion of GnRH is depressed in mid childhood, increasing with the onset of puberty. Follicle-stimulating hormone (FSH) secretion is high in the infant female, declines, and increases again at puberty. Luteinizing hormone (LH) secretion is modest in infancy, is minimal in childhood, and increases significantly during adolescence. (Root AW: Puberty in the female: Normal and aberrant. In Wallach EE, Zacur HA [eds]: Reproductive Medicine and Surgery, p 100. St Louis, CV Mosby, 1994.) |
The contribution of body fat, lean body mass, and body mass index to the onset of puberty or age of menarche is also a controversial topic.71, 72, 73 Frisch’s71 critical weight theory does not explain findings in all patients. There is great variation, making absolute thresholds unreliable, but there does seem to be a trend toward earlier puberty in moderately overweight girls and delayed puberty in severely underweight or malnourished or obese girls. Anorexia nervosa patients and intensively trained athletes can experience delayed (or arrested) puberty and are found to exhibit decreased gonadotropins, dampened gonadotropic pulsatility, and decreased levels of sex steroids. Intertwined and often indistinguishable are the roles of diet, exercise, and psychological stress, which all are apparently capable, in their extremes, of affecting the age of puberty onset.53, 59 Closely linked is the finding of increased cortisol in amenorrheic athletes,53 but in terms of a critical role in triggering normal puberty, cortisol levels do not change dramatically with puberty.
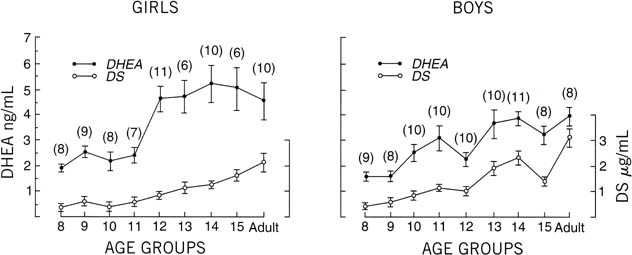
There is a progressive increase in circulating levels of DHEA and DHEA-S with sexual maturation, but only speculation exists as to a specific role of ACTH or adrenal androgens in the onset of puberty. Even the exact impetus for adrenarche remains a mystery. The chronologic alteration in circulating DHEA and DHEA-S levels has been addressed by Hopper and Yen,74 who evaluated boys and girls and adult subjects. Their research identified a progressive and parallel increase in serum DHEA and DHEA-S in boys and men. Their work identified an earlier rise for DHEA (age 13) than for DHEA-S (age 14) in boys. In addition, from the age of 8 through adulthood (in boys), there was a 2.6-fold increase in circulating levels of DHEA and a 7.7-fold increase in DHEA-S. In girls, this parallel rise in DHEA and DHEA-S was not as apparent; however, there was an abrupt increase in DHEA levels between 11 and 12 years of age. These researchers hypothesized that adrenal androgens may play a role in the “initiating factor for the central nervous system program” related to onset of pubertal development. The change in sensitivity to negative feedback at the level of the pituitary may be representative of “steroid modulation” at this central level, which is associated with increased pituitary hormone output and the related events regarding sexual maturation (Fig. 14).
{kind=link}
Under normal pubertal developmental circumstances, adrenarche and gonadarche (gonadal maturation) are closely linked. In pathologic situations, there can be a wide disarray, however, with respect to each of these entities.
Any excitement over a potential onset role of endogenous opiates, which rise during stress and may suppress GnRH or LH, was dampened with the discovery that naloxone (opiate antagonist) does not affect the initial peripubertal sleep-associated LH pulsatility.70, 74 Sex steroids themselves are unlikely culprits in the puberty trigger given that agonadal and gonadal dysgenesis patients experience comparable increased LH pulsatility patterns with puberty onset.58
No convincing role or consistent relationship has been identified for activin, inhibin, prolactin, or melatonin (with or without pineal gland), the last-mentioned being the most controversial. Because of its nocturnal secretory predominance and because it is produced by the pineal gland, which has been implicated in the pubertal onset of seasonal breeding animals, melatonin is a reasonable candidate in the mechanism of puberty control.75 It has been hypothesized that declining levels of melatonin could result in increased GnRH secretion during puberty. To date, however, no obvious relationship between melatonin blood levels and the onset and progression of normal puberty in girls has been found. With this concept in mind, research involving evaluation of plasma melatonin rhythm in humans, primarily from 24-hour serum sampling, has implicated the role of melatonin in disorders of the hypothalamic-pituitary-gonadal axis. Specifically, delayed puberty, precocious puberty, and hypothalamic amenorrhea all seem to have a direct link to altered plasma melatonin profiles (Table 6).76 Under normal circumstances, however, melatonin has less of an active role with respect to pubertal onset.77
Table 6. Studies of plasma melatonin in human puberty: normal subjects
Results |
||||
Investigators |
Subjects |
Sampling time |
No change with puberty |
Change present with puberty |
Daytime concentrations |
||||
Silman et al (1979) |
51 healthy boys and girls, 11.5–14 years; all stages of puberty |
Single sample: 1100–1300 |
No changes in plasma melatonin in girls |
Boys in stage 1 have higher plasma melatonin than all other stages |
Lenko et al (1982) |
79 girls, 83 boys, healthy, 9–16 years; all stages of puberty |
Single sample: Girls 0800–1100 Boys 1300–1500 |
No changes in plasma melatonin by sex or pubertal stage |
|
Nocturnal concentrations or 12–24-hr profiles |
||||
Arendt (1978) |
Normal children; six prepubertal, three pubertal |
Two samples: 1200, 2400 |
No change in plasma melatonin between pubertal and prepubertal children |
|
Fevre et al (1978) |
Four normal pubertal boys, 12–17 years |
Multiple samples: 20-min intervals for 24 hr |
Nocturnal increase in LH and melatonin with a positive correlation between both hormones |
|
Ehrenkranz et al (1982) |
Boys with constitutional short stature, eight prepubertal, 9–13 years and seven pubertal, 13–15 years; five normal adult males |
Multiple samples: 3-hr intervals for 24 hr |
No difference in concentration or timing of plasma melatonin among groups |
|
Waldhauser et al (1984) |
38 boys, 20 girls, 1–18 years; hospitalized in a pediatric or otolaryngological unit |
Two samples: 0700–1000 2300–0100 |
Plasma melatonin decreases as a function of pubertal stage. Inverse correlation between nocturnal melatonin and LH concentrations |
|
Attanasio et al (1985) |
Normal children, 21 girls, 17 boys, 1–18 years hospitalized for minor illness or diagnostic evaluations; all stages of puberty |
Multiple samples: 3-hr intervals from 2100 to 0600 and single sample 1200 |
Decline in 0300 melatonin concentration with age and pubertal stage from infancy to stage 5 |
|
Cavallo (1992) |
Normal children, 5–17 years; 30 boys, 32 girls; all stages of puberty |
Multiple samples: 1-hr samples by constant withdrawal from 1800 to 0800 |
No change in duration of nocturnal melatonin surge or time of peak with puberty; wide individual variation in melatonin concentrations |
Decline in nocturnal melatonin peak with age and pubertal stage, but no significant relation with pubertal stage |
LH, luteinizing hormone.
Cavallo A: Melatonin and human puberty: Current perspectives. J Pineal Res 15:115, 1993.
The gonadotropin attenuating factor (GnSAF) appears to be one additional factor associated with the functional hypothalamic-pituitary-ovarian axis physiology. GnSAF is a nonsteroidal ovarian substance associated with the midcycle LH surge.78 Its mechanism of action is related to control of the midcycle LH surge. GnSAF antagonizes the sensitizing effect of estradiol on the pituitary gland.
Other factors that appear to be related to regulation of puberty include GPR54 and KiSS-1. GPR54 and its cognate ligand, kisspeptin appear to be localized in the hypothalamus.79 KiSS-1 mRNA has been detected in the hypothalamus. In addition, GPR54 is identified on histologic assessment in the hypothalamus. Kisseptin is associated with Gonadotropic releasing hormone (GnRH) associated with the release of the gonadotropins FSH and LH. Of note is the reported increase in hypothalamic KiSS-1 and GPR54 mRNA at the time of puberty. Kuohung et al have noted that the way the mechanism of action of kisspeptin activation of GnRH release and the pathway related to onset of puberty have yet to be finalized.79 Potential exists with regard to how modulators of GPR54, which would include kisspeptin, may be utilized in cancer therapy as well as reproductive healthcare in general.
Leptin is a hormone secreted by adipocytes. It plays an integral role in energy signaling. It appears to play a permissive role in initiation of onset of puberty.80 Leptin is a 167 amino acid protein and is the product of the ob gene. It is a member of the cytokine family and is found in free in the circulation. Leptin is primarily the product of white adipose tissue and appears to be secreted in a pulsatile manner with diurnal variation.81 In observational studies there appears to be an increase in circulating levels of leptin immediately preceding onset of puberty.82 Obese children have high leptin levels and may be related to earlier onset of puberty.
In boys approaching puberty, an abrupt fall in the concentration of melatonin with advancing development suggests that this hormone may play an integral role in the physiologic onset of puberty in boys (Fig. 15). It is hypothesized that melatonin acts directly on the gonads to suppress function.83 Overall the effects seem to be predominant in boys compared with girls.
|
Fig. 15. Genitalia stage. Mean concentration (calculated from log transformed data) of melatonin (•), follicle-stimulating hormone (FSH) (▪), luteinizing hormone (LH) ( |
With respect to pubertal aberration and melatonin secretion, there seem to be higher melatonin concentrations in association with delayed puberty.84 From the opposite perspective (i.e. precocious puberty), there is a lower day-night increment in melatonin levels compared with controls. The overall conclusion is that lower circulating melatonin levels are associated with precocious puberty.85, 86 Most researchers would concur that the role of the human pineal gland with respect to pubertal development continues to be speculative. In large part, many of these studies are difficult to conduct because of the effects of light exposure and overall lack of understanding at the receptor level with respect to specific (i.e. direct) effects of melatonin on the reproductive system.
hGH has been implicated as having a role in the onset of puberty. After all, the increased hGH pulse amplitude noted with early puberty is secreted mainly during the first hours of sleep, as is GnRH and LH initially. hGH is a puberty promoter, as outlined earlier; however, puberty occurs in girls with hGH deficiency, making hGH an unlikely major factor in the initiation of puberty.
Many genes have been identified and are involved in reproduction. They are outlined in Table 7.
Table 7. Genes involved in reproduction and their mutation
Gene |
Chromosome location |
Mutations |
Phenotype |
GnRH |
8p11 |
None |
? IHH |
KAL |
Xp22.3 |
Inactivating |
IHH and anosmia |
DAX-1 |
Xp21 |
Inactivating |
AHC and IHH |
GnRH receptor |
4q21.2 |
Inactivating |
IHH |
α-Subunit |
6q |
None |
? IHH and hypothyroidism |
LH-β |
19q13.3 |
Inactivating |
Isolated LH deficiency |
hCG-β |
19q13.3 |
None |
? |
FSH-β |
11P13 |
Inactivating |
Isolated FSH deficiency |
LH/hCG receptor |
2p21 |
Activating |
Familial precocious puberty; no effect on females |
Inactivating |
Undermasculinization of genetic males; secondary amenorrhea in females |
||
FSH receptor |
2p21 |
Inactivating |
Premature ovarian failure in females; oligospermia in males |
Activating |
? Increased sperm |
||
GnRH, gonadotropin-releasing hormone; KAL, Kallmann syndrome; DAX-1, dosage-sensitive sex reversal. Adrenal hypoplasia congenital, critical region of X chromosome, gene 1; LH, luteinizing hormone; FSH, follicle-stimulating hormone; hCG, human chorionic gonadotropin; AHC, adrenal hypoplasia congenital; IHH, idiopathic hypogonadotropic hypogonadism.
SUMMARY
Puberty, the period of transition from childhood to adulthood, is composed of a multitude of dynamic processes involving psychosocial issues, secondary sex characteristics, and stature or body composition. Changes also occur in the reproductive organs themselves. Crucial to these events is a functional HPO axis and a normal karyotype. The actual precipitant of puberty remains a point of further identification.
REFERENCES
Marshall W, Tanner J: Variations in patterns of pubertal changes in girls. Arch Dis Child 44:291, 1969 |
|
Rosenfield RL: Puberty and its disorders in girls. Endocrinol Metab Clin N Am 20:15, 1991 |
|
Wheeler MD: Physical changes of puberty. Endocrinol Metab Clin N Am 20:1, 1991 |
|
Lee PA: Normal ages of pubertal events among American males and females. J Adolesc Health Care 1:26, 1980 |
|
Barnes HV: Physical growth and development during puberty. Med Clin North Am 59:1305, 1975 |
|
Taranger J, Engstrom I, Lichtenstein H, Svennberg-Redegren I: Somatic pubertal development. Acta Paediatr Scand Suppl 258:121, 1976 |
|
Karlberg P, Engstrom I, Lichtenstein H, et al: Biological interpretation of anthropometric growth data. Proceedings of the 13th International Congress on Pediatrics, Vienna, Austria 1971 |
|
Marshall WA, Tanner JM: Variations in the pattern of pubertal changes in boys. Arch Dis Child 45:13, 1970 |
|
Brook CGD, Hindmarsh PC: The somatotropic axis in puberty. Endocrinol Metab Clin N Am 21:767, 1992 |
|
Rogol AD: Growth and growth hormone secretion at puberty: The role of gonadal steroid hormones. Acta Paediatr 383(suppl):15, 1992 |
|
Mauras N, Blizzard RM, Link K, et al: Augmentation of growth hormone secretion during puberty: Evidence for a pulse amplitude-modulated phenomenon. J Clin Endocrinol Metab 64:596, 1987 |
|
Delemarre-Van De Waal HA, Wennink JMB, Odink RJH: Gonadotropin and growth hormone secretion throughout puberty. Acta Paediatr Scand 372(suppl):26, 1991 |
|
Ogilvy-Stuart AL, Shalet SM: Growth hormone and puberty. J Endocrinol 135:405, 1992 |
|
Rose SR, Municchi G, Barnes KM, et al: Spontaneous growth hormone secretion increases during puberty in normal girls and boys. J Clin Endocrinol Metab 73:428, 1991 |
|
Katz E, Ricciarelli E, Adashi EY: The potential relevance of growth hormone to female reproductive physiology and pathophysiology. Fertil Steril 59:8, 1993 |
|
Attie KM, Ramirez NR, Conte FA, et al: The pubertal growth spurt in eight patients with true precocious puberty and growth hormone deficiency: Evidence for a direct role of sex steroids. J Clin Endocrinol Metab 71:975, 1990 |
|
Bourguignon JP: Review: Linear growth as a function of age at onset of puberty and sex steroid dosage: Therapeutic implications. Endocrinol Rev 9:467, 1988 |
|
Morales AJ, Holden JP, Murphy AA: Pediatric and adolescent gynecologic endocrinology. Curr Opin Obstet Gynecol 4:860, 1992 |
|
Massa G, Maes M, Heinrichs C, et al: Influence of spontaneous or induced puberty on the growth promoting effect of treatment with growth hormone in girls with Turner’s syndrome. Clin Endocrinol 38:253, 1993 |
|
LeRoith D, Adams M, Werner H, Roberts C: Insulinlike growth factors and their receptors as growth regulators in normal physiology and pathologic states. Trends Endocrinol Metab 2:134, 1991 |
|
Clemmons D: Insulin-like growth factor binding proteins. Trends Endocrinol Metab 1:412, 1990 |
|
Nobels F, Dewailly D: Puberty and polycystic ovarian syndrome: The insulin/insulin-like growth factor 1 hypothesis. Fertil Steril 58:655, 1992 |
|
Ernst M, Health JK, Rodan GA: Estradiol effects on proliferation, messenger ribonucleic acid for collagen and insulin-like growth factor-I, and parathyroid hormone-stimulated adenylate cyclase activity in osteoblastic cells from calvariae and long bones. Endocrinology 125:825, 1989 |
|
Corvol MT, Carroscosa A, Tsagris L, et al: Evidence for a direct in vitro action of sex steroids on rabbit cartilage cells during skeletal growth: Influence of age and sex. Endocrinology 120:1422, 1987 |
|
Tamborlane WV, Amiel SA: Diabetes control and growth hormone: New insights. Growth Genet Horm 2:5, 1986 |
|
Kline KO, Barren J, Colli MJ, et al: Estrogen levels in childhood determined by an ultra-sensitive recombinant cell bioassay. J Clin Invest 94:2475, 1994 |
|
Albanese A, Stanhope R: Growth hormone deficiency throughout puberty. J Endocrinol Invest 15:777, 1992 |
|
Ritzen Ritzén EM: Pubertal growth in genetic disorders of sex hormone action and secretion. Acta Paediatr Suppl 383:22, 1992 |
|
Danziger Y, Mukamel M, Zeharia A, et al: Stunting of growth in anorexia nervosa during the prepubertal and pubertal period. Isr J Med Sci 30:581, 1994 |
|
Brinch M, Manthorpe T: Short stature as a possible etiological factor in anorexia nervosa. Acta Psychiatr Scand 76:328, 1987 |
|
Theintz GE, Howald H, Weiss U, Sizonenko PC: Evidence for a reduction of growth potential in adolescent female gymnasts. J Pediatr 122:306, 1993 |
|
Theintz GE: Endocrine adaptation to intensive physical training during growth. Clin Endocrinol 41:267, 1994 |
|
Zacharias L, Rand WM: Development and growth in American girls: The statistics of menarche. Obstet Gynecol 31:325, 1976 |
|
Greulich WW, Pyle SI: Radiographic Atlas of Skeletal Development of the Hand and Wrist. 2nd ed.. Stanford, Stanford University Press, 1959 |
|
Tanner JM: The development of the reproductive system. Growth at Adolescence. p 23, Oxford, Blackwell Scientific, 1962 |
|
Ivarsson S-A, Nilsson KO, Persson P-H: Ultrasonography of the pelvic organs in prepubertal and postpubertal girls. Arch Dis Child 58:352, 1983 |
|
Bridges NA, Cooke A, Healy MJ, et al: Standards for ovarian volume in childhood and puberty. Fertil Steril 60:456, 1993 |
|
Saxton DW, Farquhar CM, Rae T, et al: Accuracy of ultrasound measurements of female pelvic organs. Br J Obstet Gynaecol 97:695, 1990 |
|
Styne DM: Puberty and its disorders in boys. Endocrinol Metab Clin N Am 20:43, 1991 |
|
Harlan WR, Grillo GP, Cornoni-Huntley J, et al: Secondary sex characteristics of boys 12 to 17 years of age: The US health examination survey. J Pediatr 95:293, 1979 |
|
Croft CA, Asmussen L: A developmental approach to sexuality education: Implications for medical practice. J Adolesc Health 14:109, 1993 |
|
Lipsius SH: Normal sexual development of children: Physician roles in bridging gaps in parent-child communication. Md Med J 41:401, 1992 |
|
Ogundari JT: Somatic deviations in adolescence: Reactions and adjustments. Adolescence 20:179, 1985 |
|
Tanner JM: Sequence, tempo and individual variations in the development of boys and girls aged 12 to 16. In Grinder RE (ed): Studies in Adolescence. New York, Macmillan, 1975 |
|
Rabinovici J: The differential effects of FSH and LH on the human ovary. Baillieres Clin Obstet Gynaecol 7:263, 1993 |
|
Beck-Peccoz P, Padmanabhan V, Baggiani AM, et al: Maturation of hypothalamic-pituitary-gonadal function in normal human fetuses: Circulating levels of gonadotropins, their common alpha-subunit and free testosterone, and discrepancy between immunological and biological activities of circulating follicle-stimulating hormone. J Clin Endocrinol Metab 73:525, 1991 |
|
Rosenfield RL: The ovary and female sexual maturation. In Kaplan SA (ed): Clinical Pediatric Endocrinology. pp 259, 324 Philadelphia, WB Saunders, 1990 |
|
Kulin HE: Editorial: Puberty: When? J Clin Endocrinol Metab 76:24, 1993 |
|
Yen SS, Apter D, Bützow T, Laughlin GA: Gonadotrophin releasing hormone pulse generator activity before and during sexual maturation in girls: New insights. Hum Reprod 8:66, 1993 |
|
Apter D, Butzow L, Laughlin G, Yen S: Gonadotropin-releasing hormone pulse generator activity during pubertal transition in girls: Pulsatile and diurnal patterns of circulating gonadotropins. J Clin Endocrinol Metab 76:940, 1993 |
|
Apter D: Ultrasensitive new immunoassays for gonadotropins in the evaluation of puberty. Curr Opin Pediatr 5:481, 1993 |
|
Styne DM: Physiology of puberty. Horm Res 41(suppl 2):3, 1994 |
|
Carpenter SE: Psychosocial menstrual disorders: Stress, exercise and diet’s effect on the menstrual cycle. Curr Opin Obstet Gynecol 6:536, 1994 |
|
Marshall JC, Kelch RP: Gonadotropin-releasing hormone role of pulsatile secretion in the regulation of reproduction. N Engl J Med 315:1459, 1986 |
|
Genazzani AD, Petraglia F, Gastaldi M, et al: Evidence suggesting an additional control mechanism regulating episodic secretion of luteinizing hormone and follicle stimulating hormone in pre-pubertal children and post-menopausal women. Hum Reprod 9:1807, 1994 |
|
Bridges NA, Matthews DR, Hindmarsh PC, Brook CGD: Changes in gonadotropin secretion during childhood and puberty. J Endocrinol 141:169, 1994 |
|
Bernstein L, Pike MC, Ross PK, Henderson BE: Age at menarche and estrogen concentrations of adult women. Cancer Causes Control 2:221, 1991 |
|
Conte FA, Grumbach MM, Kaplan SL: A diphasic pattern of gonadotropin secretion in patients with the syndrome of gonadal dysgenesis. J Clin Endocrinol Metab 40:670, 1975 |
|
Herman-Bonert VS, Braunstein GD: Gonadotropin secretory abnormalities. Endocrinol Metab Clin N Am 20:519, 1991 |
|
Mansfield MJ, Emans SJ: Review: Anorexia nervosa, athletics, and amenorrhea. Pediatr Clin North Am 36:533, 1989 |
|
Weiner H: Psychoendocrinology of anorexia nervosa. Psychiatr Clin North Am 12:187, 1989 |
|
Eisenberg E: Toward an understanding of reproductive function in anorexia nervosa. Fertil Steril 36:543, 1981 |
|
Sheikholislam BM, Stempfel RS: Hereditary isolated somatotropin deficiency: Effects of human growth hormone administration. Pediatrics 49:362, 1972 |
|
Kol S, Adashi EY: Intraovarian factors regulating ovarian function. Curr Opin Obstet Gynecol 7:209, 1995 |
|
Dunger DB, Perkins JA, Jowett TP, et al: A longitudinal study of total and free thyroid hormones and thyroxine binding globulin during normal puberty. Acta Endocrinol (Copenh) 123:305, 1990 |
|
Holder AT, Wallis M: Actions of growth hormone, prolactin and thyroxine on serum somatomedin-like activity and growth in hypopituitary dwarf mice. J Endocrinol 4:223, 1977 |
|
Solomon J, Greep RO: The effect of alterations in thyroid function on the primary growth hormone content and acidophil cytology. Endocrinology 65:158, 1959 |
|
Fisher DA, Sach J, Oddie TH, et al: Serum T4, TBG, T3 uptake, T3, reverse T3 and TSH concentrations in children 1 to 15 years of age. J Clin Endocrinol Metab 45:191, 1977 |
|
Bellastella A, Crriscuolo T, Mango A, et al: Circannual rhythms of plasma growth hormone, thyrotropin and thyroid hormones in pre-puberty. Clin Endocrinol 20:531, 1984 |
|
Root AW: Puberty in the female: Normal and aberrant. In Wallach EE, Zacur HA (eds): Reproductive Medicine and Surgery. pp 79, 135 St Louis, CV Mosby, 1995 |
|
Frisch RE: Fatness and fertility. Sci Am 258:88, 1988 |
|
Scott EC, Johnston FE: Review: Critical fat, menarche, and the maintenance of menstrual cycles: A critical review. J Adolesc Health Care 2:249, 1982 |
|
de Ridder CM, Bruning PF, Zonderland ML, et al: Body fat mass, body fat distribution, and plasma hormones in early puberty in females. J Clin Endocrinol Metab 70:888, 1990 |
|
Hopper BR, Yen SSC: Circulating concentrations of dehydroepiandrosterone and dehydroepiandrosterone sulfate during puberty. J Clin Endocrinol Metab 40:459, 1975 |
|
Cavallo A: Melatonin and human puberty: Current perspectives. J Pineal Res 15:115, 1993 |
|
Cavallo A, Zhou XH: LHRH test in the assessment of puberty in normal children. Horm Res 41:10, 1994 |
|
Silman RE, Leone RN, Cooper RJL, Preece MA: Melatonin, the pineal gland and puberty. Nature 282:301, 1979 |
|
Messinis I. From menarche to regular menstruation: endocrinological background. Ann NY Acad Sce 2006;1092:49-56 |
|
Kuohung W, Kaiser U. GPR54 and KiSS-1: role in the regulation of puberty and reproduction. Rev Endocr Metab Disord 2006;7(4):257-63. |
|
Kelesidis T, Mantzoros C. The emerging role of leptin in humans Peditr Endocrinol Rev 2006;3:239-48 |
|
Chan J, Mantzoros C. Role of leptin in energy-deprivation states: normal human physiology and clinical implications for hypothalamic amenorohoea and anorexia nervosa, Lancer 2005;366:74-85 |
|
Rogol A. Leptin and puberty J Clin Endocrinol Metab 1998;83:1089-90 |
|
MacPhee AA, Cole FE, Rice BF: The effect of melatonin on steroidogenesis by the human ovary in vitro. J Clin Endocrinol Metab 40:688, 1975 |
|
Cohen HN, Hay LD, Annesley TM, et al: Serum immunoreactive melatonin in boys with delayed puberty. Clin Endocrinol 17:517, 1982 |
|
Attansio A, Borrelli P, Gupta D: Circadian rhythms in serum melatonin from infancy to adolescence. J Clin Endocrinol Metab 61:388, 1985 |
|
Waldhauser FG, Weiszenbacher E, Tatzer B, et al: Alterations in nocturnal serum melatonin levels in humans with growth and aging. J Clin Endocrinol Metab 66:648, 1988 |